Dominance Styles and Variability of Social Relationships in Non-Human and Human Primates

Almanac: Evolution:Evolutionary Aspects: Stars, Primates, and Religion
The continuity of social life observed between non-human primates and humans is fundamental for understanding the formation of human society in the course of evolution as well as its further social evolution. This paper aims specifically at studying dominance styles and variability of social relationships in non-human primates and humans. The statistical analysis of our primatological database has shown that the egalitarian type of social organization in primates is associated with the following traits: 1) absence or low incidence of submission demonstration; 2) permissiveness of non-subordinate behavior towards dominants; 3) low level of male leadership power (i.e., the situation when group members systematically dispute the leader); 4) permissiveness on feeding sites; 5) low intensity of contact aggression; 6) low risk of injury; 7) low risk of severe injury; 8) high level of bidirectional aggression; 9) low level of redirected aggression; 10) low level of female-female intergroup aggression; 11) low level of male-male intergroup aggression; 12) presence of formal biting; 13) support in aggression on the side of victims rather than aggressor; 14) high le vel of affiliative support; 15) low level of aggressive support; 16) high level of reconciliation after conflict; 17) low kin preferences in reconciliation; 18) low or absent kin preferences in affiliation; 19) high probability of female carrying other females infants (i.e., permissiveness in access to infants for other females, non-kin included); 20) low or absent kin effect on spatial distribution in feeding; 21) presence of social play between males; 22) high level of male affiliative interaction with juveniles; 23) high level of male-male tolerance (and late emigration of adolescent males in case of female phylopatric species); 24) high level of grooming between males; 25) high level of grooming between non-kin females; 26) absence of connection between grooming received and dominant status; 27) high female choice of reproductive partner; 28) high female status; 29) different degrees of egalitarianism within males vs. females (in some cases); 30) presence of male attachments with anestrus females; 31) absence of seasonal prevalence of reproduction. The non-egalitarian primate social systems are characterized generally by the same set of parameters with opposite values. Our further statistical tests have shown that the ‘primate’ correlation matrix for the egalitarian vs. non-egalitarian characteristics, while completely valid for the simple hunter-gatherer cultures, almost disintegrated for the medium-range societies but resurfaced again among the most complex cultures.
Keywords: ethology, evolutionary anthropology, primatology, social evolution.
Introduction
The continuity of social life observed between non-human primates and humans is fundamental for understanding the formation of human society in the course of evolution (Butovskaya and Rostovtseva 2021; Butovskaya and Fainberg 1993; Grinin et al. 2009, 2011; Muthukrishna et al. 2018; Butovskaya 2020) as well as its further social evolution. The capacity for self-recognition, purposefulness, long-term memory, prediction of others' actions, deception, the understanding of social bonds within the group – these are some, but by no means all, prerequisites of human society, those which are actually observed in extant great apes. Like many other phenomena of human life, certain aspects of social relationships and culture can be explained from the perspective of natural sciences (Daly and Wilson 1988a, 1988b; Rodseth et al. 1991; Eibl-Eibesfeldt and Sutterlin 1992; Wrangham and Peterson 1996; Street et al. 2017; Azumagakito et al. 2018). An important task is to disclose continuity between primate social structures and human society and to solve certain problems concerning the biological roots of human social institutes and properties such as systems for transferring social information, systems of kinship, marriage and social stratification (Betzig 1997; Butovskaya and Fainberg 1993; Butovskaya 1999a, 1999b; Lancaster 1997; Blurton Jones et al. 1999; Hrdy and Burkart 2020; Butovskaya and Rostovtseva 2021). The present paper, based on evidence from non-human and human studies, attempts to demonstrate some basic features of social structure and in-group and out-group social relationships observed in non-human and human primates. The special attention will be paid to comparative analysis of social hierarchy types and their relevance to ecology and phylogeny.
Modern humans are known to possess the highest inter-population differences in social structure and types of hierarchical relationships compared to non-human primates. With respect to the dominance relations the two main types of social organization, known among the non-human primates, are normally denoted as (1) ‘egalitarian’ or ‘tolerant’ and (2) ‘non-egalitarian’ or ‘despotic’ (Verencamp 1983; van Schaik 1989; Butovskaya 1993; Thierry et al. 1990; Matsumura 1999; Zannella et al. 2017). Currently, these differences can also be described in terms of the degree of hierarchical steepness (Balasubramaniam et al. 2012; Butovskaya 2020; Ogawa 2020).
Is it possible to find similar types of social organization among humans? It is well known that the terms ‘egalitarian’ and ‘non-egalitarian’ are widely used in sociocultural anthropology to denote certain types of the human social organization. However, in most cases the respective terminological tradition ascends to the legacy of Fried (1967), and Service (1962[1971]), whereby the ‘egalitarian societies’ are understood as ‘a society in which all persons of a given age-sex category have equal access to economic resources, power, and prestige’ (Ember and Ember 1999: 505; for a similar understanding of the egalitarianism vs. non-egalitarianism dichotomy see e.g. Haviland 1991: 515; Bailey and Peoples 1999: 174–178, etc.).
The term ‘egalitarian’ itself appears to have been borrowed by primatologists from cultural anthropology, however, now it is understood as dominance style characterized by the set of following principal behavioral patterns: less severe aggression, greater symmetry in contests, higher rate of affiliation, higher rate of peaceful post-conflict encounters (reconciliation), less effect of kinship on social interactions compared to social systems with ‘despotic’ hierarchical relations (Matsumura 1999). As will be shown below, it is possible to add some more parameters to this list. The definition which we will use in this paper, is the one defining the egalitarian communities as those characterized by ‘weakly linear or shallow dominance relations’ (Matsumura 1999: 24; originally proposed by van Schaik 1989). In addition, it is important to realize that there is no sharp distinction between these two types of social relations and species should be rather distributed along the scale ranging from maximal expressions of ‘egalitarianism’ at one end and maximum expression of ‘despotism’ at the other.
It is clear that the above-mentioned ‘cultural-anthropological’ definition does not correspond quite well to van Schaik's definition. A human society ‘in which all persons of a given age-sex category have equal access to economic resources, power, and prestige’ might be characterized by highly lineal and rigid dominance relations (as is observed, e.g. among the Australian Aborigines [Artemova 1987, 1989, 1991, 1993, 2000, 2003]), whereas a society with marked economic inequality may still be characterized by ‘weakly linear or shallow dominance relations’, as was observed, for example, among the citizens of Classical Athens (Berent 1994, 1996, 1998, 2000, 2004, 2006) or in the modern cultures of the West.
Hence, we suppose it would be more productive to apply the perspective developed by Woodburn (1972, 1979, 1980, 1982, 1988a, 1988b) and Artemova (1987, 1989, 1991, 1993, 2000, 2009, 2016, 2020) who understand the ‘egalitarianism’ just along the lines described above for the primatological field. Though they have developed this understanding independently, they have virtually shown the high applicability of the ‘primatological’ notion of egalitarianism/non-egalitarianism to the human societies. What is more, such an understanding makes it perfectly possible to compare the human and non-human primate societies; that is why we employ just this understanding in the present paper.
Socioecology and Social Complexity: Female-Bonded
and Non-Female-Bonded Societies
Social interactions in primates are supposed to be strongly influenced by dominance and kinship. To understand the reasons of species and even population differences in the types of hierarchies, dispersal patterns and relations within and between sexes, a number of hypotheses have been proposed. One group of such hypotheses is based on the idea of positive connections between ecology and sociality (Wrangham 1980; van Schaik 1989; Kappeler 2019; Moscovice et al. 2020), the other – focuses on the role of phylogenetic inertia in the development of certain models of social organization and residence strategies (Matsumura 1999; Strier 1999; Thierry 1999; Balasubramaniam et al. 2018). The latter is not surprising given the fact that social system is an outcome of the social behavior, and thus, of group members' relationships (Hinde 1983).
Are these differences connected somehow to socioecology? To answer this question, let us first address the non-human primate data. Field primatology seems to give some general answer. According to the socioecological paradigm, the degree of complexity of social relationships and social mechanisms aimed at preventing social tension depend on the ecological context in which the species exists (Wrangham 1980; van Schaik 1989; Amici et al. 2018; Vermande and Sterck 2020). The species, then, is regarded as one of the components of a local ecosystem, whereas social relationships are viewed as factors optimizing the adaptation of groups within this species to the respective ecosystem.
Two basic hypotheses concerning the reasons underlying group formation and the maintenance of in-group cohesion were proposed. The first one focuses on the need to form cohesive groups in order to successfully compete for food resources with the conspecifics on the intergroup level (Wrangham 1980). In addition, correlation has been established between terrestriality, food preferences and group size (Clutton-Brock and Harvey 1977; van Schaik 1989). According to Dunbar (1988), terrestrial frugivores and generalized omnivores tend to form large foraging groups with differentiated competitive relationships among females. Group size can also depend on the environment. Observations of chimpanzees in different environmental settings have revealed a tendency to a higher level of cohesiveness in dry and open environments (Mt. Assirik, Senegal, see Tutin et al. 1983). Larger, mixed parties with adult males are reportedly more common in non-forested habitats than are solitary individuals or groups without males. In highly fragmented habitats adolescent females do not leave the natal group, that contradicts their most female conspecifics (Sugiyama 1989). According to the second hypothesis, high predator pressure is sufficient for the selection favoring a gregarious way of life (van Schaik and van Hooff 1983). Both predator pressure and between-group competition hypotheses agree that the differences between species should exist with respect to within-group social relationships of females. Thus, the level of within-group competition reflects the pattern of resource distribution and the quality of these resources (van Noordwijk and van Schaik 1987; van Schaik 1989).
Mostly social relationships are the consequences of ecological pressures on individuals. Sociality has obvious impact on individual's fitness in primates (Ostner and Schuelke 2018). Social behavior is aimed at enhancing inclusive fitness which is different in males and females (this rule remains stable in human species). Indeed, it was found that reproductive success of males and females depended on different factors. While food is the main factor limiting females' reproductive success, the males' reproductive success is supposedly limited by the access to females (Wrangham 1980). No wonder that in the light of this paradigm female relationships were estimated as a keystone of social organization. Between-group contest is the primary stimulus for female bonding because its effect on the fitness of females is supposed to outweigh the effect of within-group competition (Butovskaya 1999a; Emery Thompson 2019). In addition, females may form groups to reduce the risk of predation and in the process of group formation, within-group competition for food may arise (van Schaik 1989). Wherever food is easily monopolized, within-group competition is of the contest type (long-tailed macaques, rhesus macaques) and female relationships are more despotic and nepotistic (kin-oriented), resulting in a female-bonded (matrilineal) group structures (van Noordwijk and van Schaik 1987; van Schaik 1989). In situations where food resources are small and dispersed, competition is of the scramble type (Saimiri spp., Mitchell et al. 1991), and non-female-bonded groups are likely to emerge. Competition between females is virtually absent where resources are abundant and distributed over large patches (Presbytes thomasi, Sterck et al. 1997). In such cases, female relationships will be highly egalitarian and dispersive and a non-female-bonded group structure is highly probable.
Recent studies of greater diversity of primate taxa, however, have shifted the initial idea about the ‘typical’ primate social structure based on female kin bonding and male dispersal (Moore 1996; Strier 1999). Consistent with this new paradigm is an idea of F. Lee (1984), that the absence of female kin bonding among hominoids is a derived condition, reflecting the monophiletic origin of great apes and reduction of predation pressure with the general increase in their body size. It is also suggested that high costs of feeding competition due to specialization on ripe fruit diets preclude chimpanzees and orangutans from forming cohesive female bonded groups because gorilla rely mainly on low- quality diet and there is no necessity to compete for food resources. No wonder that females of this species do not bond with each other but develop strong long-time bonds with males (Wrangham 1980; Schaik 1989). Unlike in the Old World primates, female bonded groups are rare in the New World primates (Strier 1999). They are formed mainly because the effect of within-group contest is generally more important than that of between-group contest. Many non-female-bonded species are folivorous (Gorilla gorilla beringei, Watts 1994) while most female-bonded ones are frugivorous (Macaca spp., Schaik 1989; Butovskaya 1993). In situations where animals feed basically on large fruit trees and supplement their diet with other types of food, within-group competition may be low but female-bonded structure is still preserved (Tonkeana macaques). When the likelihood of between-group competition is high, a female resident pattern can be expected (Erythrocebus patas, Chism and Rowell 1986).
Explanation of the evolution of female grouping in primates demands understanding feedback connections in the evolution of social relationships (Sterck et al. 1997). One of these catalyzing stimuli is the infanticidal strategy of males, which must certainly have affected female strategies in species where infants were endangered. The risk of infanticide can promote the formation of multi-female, multi-male groups (Manguette et al. 2019). This is due to the fact that within such social structure females can make paternity less certain (by not displaying external signs of ovulation, by promiscuous mating, etc.) thus ensuring a multi-male protection for their infants. Infanticide risk may be the fundamental reason for grouping in situations where predation risk, within-group contest and between-group contest are low (Ibid.). The comparison of congeneric species (e.g., those of the genus Macaca) has revealed that patterns of coping with social tension are less deve (loped in more arboreal species compared with more terrestrial ones (Macaca fascicularis compared to M. arctoides and M. tonkeana). This is due to a greater importance of sociality for the survival of single individuals in terrestrial species where group life provides a defense against predators and an access to resources under competition with conspecific groups. However, recent data on white-faced capuchin monkeys revealed that new alpha males often kill young infants sired by other males, and infants of highly ranking females are at greater risk of dying compared to less central females (Kalbitzer et al. 2017). Hence, there may be a trade-off between benefits of higher rank in terms of food access and increasing likelihood of their infants mortality due infanticide.
Van Schaik's model mentioned above mainly sought to explain the reasons underlying group formation in females. Later, an attempt was made to predict social relationships in males (van Hooff and van Schaik 1994). Since females, who are the limiting resource for males, cannot be easily shared, it was suggested that cooperation among males is less common and mainly takes the form of reciprocal altruism and cooperative alliances. As it will be demonstrated in this paper later, this factor is important for structuring the social relationships in human societies either.
Group Life History and Social Organization
In contrast to widespread ideas about the crucial role of ecology, contemporary field data suggest that the knowledge about ecological variables are not totally sufficient to explain the social system and dispersal patterns both in the New World and the Old World primates. Phylogenetic constraints in combination with reproductive, demographic and social constraints should be taken into account to understand the connections between foraging strategies, dispersal patterns and male-female social relationships in primates (Lee 1994; Butovskaya and Kozintsev 1996a; Strier 1999; Matsumura 1999). The prerequisite of an efficient social structure is a positive balance between individual cost and the benefit which sociality provides to single individuals. This balance may vary across populations, ultimately resulting in the emergence of between-population differences in dominance style and, respectively, in various behavioral models aimed at preventing social tension and removing its effects.
In a number of cases group differences in social organization can hardly be attributed to feeding ecology or population density. The problem that arises is that of the individual components in the formation of a social style and tension regulating strategies. Relationships within a group largely depend on group history and the characteristics of single individuals (Datta 1989; Butovskaya 1995). Our data on two groups of long-tailed macaques may serve a good model. Differences in dominance style that we have found between two groups of M. fascicularis approach inter-specific differences in magnitude. The two groups were kept in cages under identical conditions. Their diet was identical as well. The groups consisted of eight and ten adult animals, respectively, and each one included an alpha male. The crucial difference was that one of the groups (group H) consisted of animals which were high-ranking by birth while another one (group L) comprised only those whose mothers were low-ranking (Butovskaya and Kozintsev 1996b).
Values of Landau's index indicated that the hierarchical structure in both groups was moderately linear. However, nearly all parameters of aggressive behavior were significantly higher in the group L, with the sole exception of injury rate. Reconciliation in the group H was very rare and victims seldom redirected aggression to other individuals. They rarely sought consolation from the third parties. The alpha male in the group H was virtually the only animal who comforted the victims after aggression.
Female aggressors in the group L initiated reconciliation seven times more often than did their counterparts in the group H. In the group L, victims sought contacts with their friends, who soothed them, nine times more actively than did victims in the group H. While ritual biting was fairly common in the group L, it was almost never practiced in the group H (Ibid.). In terms of dominance style, the group H may be described as a community of a despotic type and group L as an egalitarian one. The two groups can be regarded as a model for the evolution of various dominance styles and various mechanisms of coping with social tension under identical ecological conditions (Ibid.). Thus, different groups within the same species may display marked differences in dominance style and can use various ways of regulating social equilibrium not only due to differences in ecological conditions but as the consequence of unique group history, and unique individualities of group members.
The Phylogeny and Similarity in Social Organization
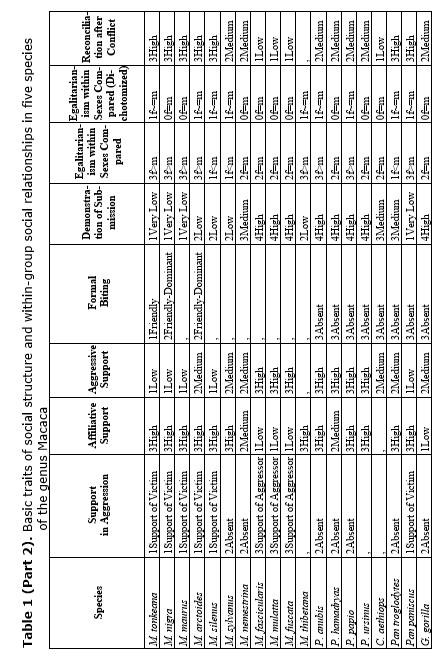
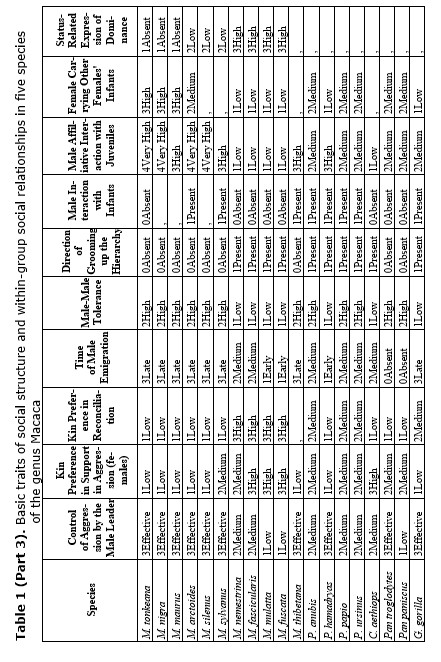
Sometimes, the variation of social strategies cannot be ascribed either to ecological factors or to group history and phylogeny may be ‘a major determinant of social relationships’ (Thierry et al. 1990). This hypothesis seems to be correct for the explanation of main patterns of social organization in the genus Macaca (Thierry 1999). Thus, according to proposed 4-grade scale of social organization, dominance styles and patterns of social activity are distributed from rigid to relaxed egalitarian. Macaca mulatta and M. fuscata belongs to grade 1, being the most nepotistic, with largely asymmetric and dictatorial relationships (Butovskaya 1993; Thierry 1985, 1999). Low-ranking individuals always keep an eye on the alpha male (this is the essence of the attention structure phenomenon), try to please him and avoid direct competition with him for food or sexual partners. They must demonstrate submission to the alpha male in order not to be attacked by him. Grade 2 represented by Macaca fascicularis and M. nemestrina demonstrate the great similarity in social patterns to grade 1, but all traits are less extreme. Grade 3, includes M. arctoides, M. assamensis, M. radiata, M. thibetana, etc.) is similar to grade 4. In this grade group social relationships are mild and high-ranking animals are interested in maintaining relationships with the subordinates (Butovskaya 1993). Grade 4 represented by Solawesi species (M. tonkeana, M. nigra) exhibit the most symmetrical and egalitarian social relations (Thierry 1985; Matsumura 1999). Both in grade 4 and 3 the overall affiliation level within the group is higher, subordinates enjoy greater freedom and can themselves initiate contacts with the dominants. A typical pattern used by the dominants to neutralize their aggressive motivation is ritual biting with a low probability of injuries.
Kinship and Dominance Style
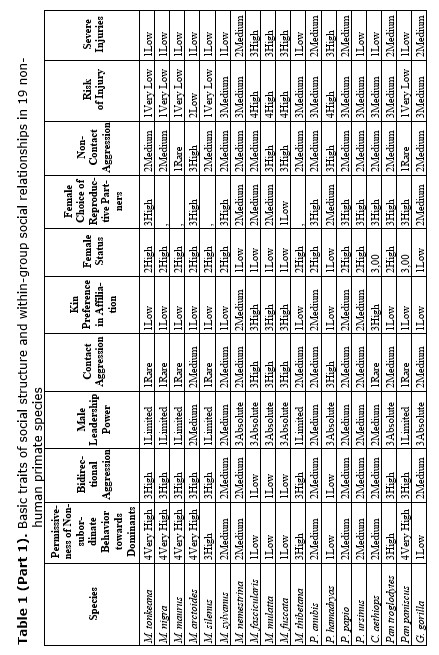
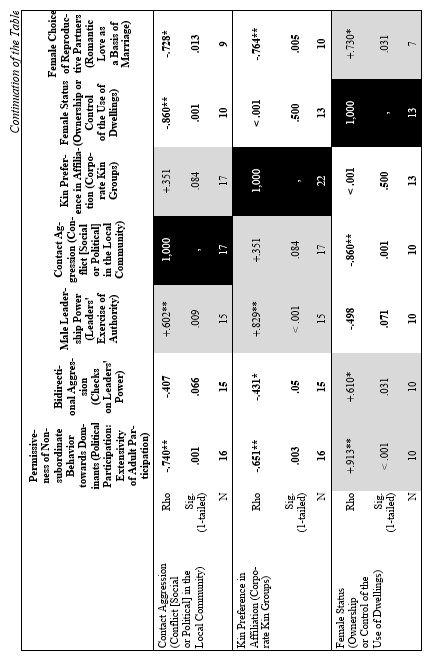
Similar dominance relations can be observed in species with different dispersal patterns and phylogenetic position. The database composed of 40 behavioral patterns was created by us to provide a detailed description of social relationships in 19 non-human primate species to estimate a set of traits characteristic for egalitarianism vs. despotism (see Table 1). The list of species contains one species of Cercopithecus, eleven species from genus Macaca, four species from genus Papio and three African ape species.
Kinship relations seem to be among the most important factors for the maintenance of group cohesion. Numerous field observations have been already demonstrated, that relatives are more predisposed to support and protect each other (see Table 1). They can cooperate in the rearing of infants (females) or protect females from alien males. It has also been shown that close attachment between relatives is based on familiarity. The familiarity factor is important for both males and females irrespective of the type of migration. Even in female-bonded macaque societies, close kinship relations between the male and its female relatives may be stable over the entire lifespan (M. arctoides, M. tonkeana) (see Table 1). Related females, if they have not emigrated, are usually more attached to each other in male-phylopatric species than the non-related females are (see Table 1). In hamadryas baboons, chimpanzees, bonobo and gorillas, close female kinship preferentially support each other in aggressive encounters (see Table 1) (Goodall 1986; Kano 1992; Watts 1992; Chalian et al. 1997; Franz 1999).
The degrees of intensity of social relations among group members, as well as the degree of their rigidity, are far from being homogeneous. It was demonstrated that in the genus Macaca, in which all species form matrilineal social structures, some positive correlation exists between the degree of despotism and the strength of kinship ties (see Table 1) (Silk 1982; Butovskaya 1993; Matsumura 1999). The general rule for this taxon can be formulated as follows: more despotic societies are more nepotistic. Altruistic behavior under such conditions is basically directed towards close kinship (mother-children, siblings, grandmother-grand-daughters). It is highly probable that altruistic behavior, having evolved in the context of close kinship, can be redirected towards other group members for instance in situations of reciprocal altruism. Egalitarianism is also probable when in-group competition for food resources is not intense. Cercopithecus aethiops and savanna baboons, represented in our sample by Papio anubis, P. ursinus and P. papio also been females bonded and quite egalitarian, while P. hamadryas is non-females bonded and non-egalitarian (see Table 1) (Dunbar 1988, 1994; Petit et al. 1997; Kummer 1971; Colmenares et al. 1994; Chalian et al. 1997; Butovskaya et al. 1998).
Recent works by a number of investigators dealing with social relationships in various macaques, baboons and African great apes species have demonstrated a high degree of coadaptation between various behavioral traits (see Table 1). It has been shown that highly intense and severe aggression (high frequency of biting and wounding) is closely related to fleeing and submission, while in situations with a low risk of injuries, high probability of reconciliation is expected (see Table 1). Asymmetry in dominance and kinship relations is in close positive correlation with asymmetry of interactions. In species with small rank differences, reconciliatory tendencies are high, inter-individual distances are minimal, aggressive interactions are largely bi-directional, affiliative interactions between group members are very common regardless of rank or relatedness between the partners (Macaca arctoides, M. tonkeana, M. silenus, M. sylvanus) (see Table 1) (Thierry 1988; Butovskaya 1993, 1995; Abegg et al. 1996; Butovskaya and Kozintsev 1996a).
In contrast, species with marked hierarchical relations display mostly unidirectional and severe aggression, the choice of affiliative partners is largely limited to kinship and group-mates of similar rank, victims and aggressors are less inclined to reconcile (see Table 1) (M. mulatta, M. fascicularis, M. fuscata, M. nemestrina) (de Waal and Luttrell 1989; Thierry 1990; Butovskaya 1993). Usually adult males are dominant over females but these species with despotic dominant relations matrilineal ties are so strong that sometimes females may jointly attack dominant male and even injure him (M. mulatta, M. fascicularis). Consequently, in these species dominant males could fail to control the in-group aggression from time to time. This became especially evident during disputes over power between matrilines (see Table 1). For example, we observed the case of severe fights between dominant and second-ranked matriline in the colony of long-tailed (the Sukhumi Primate Center), during which six animals were killed and many others were severely injured. All attempts of dominant male to stop the aggression were not effective. The conflict resulted in redistribution of power in the group: the former dominant matriline had fallen into the very bottom and became the lowest ranking, while the opponent matriline took the dominant position. The division of macaques species into egalitarian and despotic is not absolute. Generally, these species display different states of equilibrium, varying along a continuum from more egalitarian type to more despotic (Thierry 1990). A comparison of eleven macaque species on the basis of data obtained by other investigators and our own ones is presented to demonstrate this statement (see Table 1) (Thierry 1988, 1999; de Waal and Luttrell 1989; Aureli et al. 1989; Butovskaya 1993, 1995; Matsumura 1999). These data are supported by data on other Old World primate species from genus Ceropithecus, Papio, Pan and Gorilla (see Table 1).
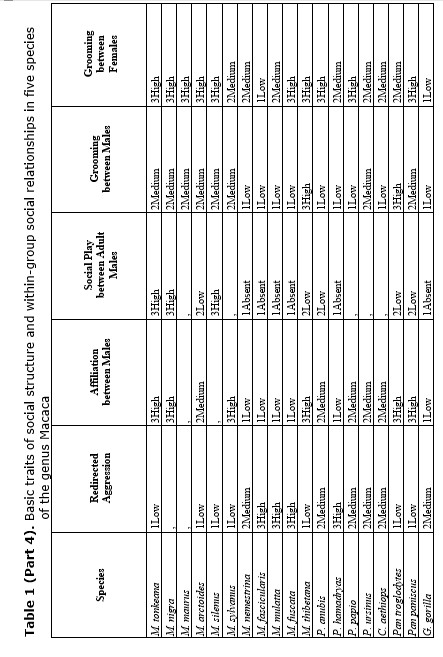
What about social relations between males in female-bonded and female-resident societies? The data revealed that close male relations in this case may be possible. M. arctoides, M. thibetana are the examples (see Table 1) (Zhao 1993; Butovskaya and Kozintsev 1996a; Matsumura 1996, 1999). In these species, although being non-related, males are highly tolerant to each other and spend much time in close proximity, they intervene in dyadic disputes among other males and frequently reconcile after conflicts (Silk 1982; Butovskaya 1993; Matsumura 1996). However, it was shown by us that male stump-tailed macaques can manipulate their affiliative preferences in favor of more profitable partners. Nevertheless, kinship ties do exist and sometimes males prefer to choose relatives for affiliation if the rank factor is excluded (see Table 1).
M. tonkeana: Thierry 1985, 1988, 1990; Thierry et al. 1990;
M. nigra: Petit and Thierry 1994a; Petit et al. 1997; Thierry 1999;
M. maurus: Matsumura 1996, 1999; Thierry 1999; van Schaik et al. 1999;
M. arctoides: Bertrand 1969; Estrada 1984; Waal and Luttrell 1989; Butovskaya 1993; Butovskaya and Kozintsev 1996a;
M. silenus: Abegg et al. 1996; Thierry 1999; van Schaik et al. 1999;
M. sylvanus: Deag 1974; Taub 1980; Kuester and Paul 1986, 1992; Paul and Kuester 1987; Aureli et al. 1994; Preuschoft et al. 1998;
M. nemestrina: Caldecott 1986a, 1986b; Judge 1991; Butovskaya 1995; van Schaik et al. 1999;
M. fascicularis: Thierry 1985, 1988; van Noordwejk and van Schaik 1987; van Schaik 1989; Aureli 1992; Aureli and van Schaik 1991; Butovskaya 1993, 1995;
M. mulatta: Thierry 1985; Waal and Luttrell 1989; Waal 1990; Waal and Yoshihara 1983; Butovskaya 1993, 1995;
M. fuscata: Aureli et al. 1993; Takahata et al. 1994; Saito 1996; Petit et al. 1997;
M. thibetana: Deng and Zhao 1987; Zhao and Deng 1988; Zhao 1993; Ogawa 1995; Matsumura 1999;
P. anubis: Sugawara 1988; Barton 1993; Barton et al. 1996; Castles and Whiten 1998a, 1998b;
P. hamadryas: Kummer 1971; Sigg 1980; Colmenares et al. 1994; Colmenares and Lozaro-Perea 1994; Chalian et al. 1997; Butovskaya et al. 1998;
P. papio: Sayfarth 1978; Petit and Thierry 1994b; Petit et al. 1997;
P. ursinus: Busse and Hamilton 1981; Hausfater et al. 1982; Noe 1986; Silk et al. 1996; Palombit et al. 1997;
C. aethiops: Cheney and Seyfarth 1986, 1989;
Pan troglodytes: Tutin et al. 1983; Goodall 1986; Nishida and Hiraiwa-Hasegawa 1987; Sugiyama 1989; Moore 1996;
Pan paniscus: Nishida and Hiraiwa-Hasegawa 1987; Susman 1987; Waal 1987; Ihobe 1992; Kano 1992; Kuroda 1984; Hohmann and Fruth 1993;
Gorilla gorilla: Harcourt 1978; Watts 1992, 1994, 1995; Sicotte 1993, 1995.
Social Hierarchy and Dominant Sex
The data from non-human primate socioecology revealed the complex interconnection between dispersal patterns and patterns of inter- and between-sex dominance relations. It has been already demonstrated that social relationships in female-resident species (macaques, savanna baboons) varied along the range from highly egalitarian on one end to highly despotic on the other. Let us now address the social relationships in male-resident species. For example, both chimpanzees and bonobos are male-phylopatric but they strikingly differ in the pattern of inter-male and inter-female relationships (see Table 1). In chimpanzees, males maintain close connections with each other. Such associations are tightly associated with formalized dominance hierarchies, and alliances are likely to change following the restructuring of male-dominance relations (Goodall 1986). Grooming is not a reflection of attachment between the male kin, but rather a social tactics to form alliances against other individuals. In bonobos, on the other hand, male dominance hierarchy is less clear-cut, males show loose associations, and alliances in aggressive conflicts are rare (Susman 1987). Moreover, females frequently dominate males in conflicts over food (see Table 1) (Kano 1992). It is suggested that only cooperation and mutual support of females enable their dominance (Franz 1999; Kano 1992). Because most of adult females in bonobo groups are usually non-kin, the only explanation of this phenomenon is that newly emigrant females practiced a special ‘social adaptation’, they established friendly bonds with older and higher ranking females. Unlike in male chimpanzees, grooming relations in female bonobos are correlated with friendly bonds and in no way grooming in this species can be explained as a pay-off for support from the side of higher-ranking animals. It means that dominant bonobo females were not groomed more than subordinate ones and dominant females even initiated more grooming down the hierarchy (see Table 1) (Franz 1999). Food sharing between the males is less common. Female-female relationships in bonobos are characterized by a high level of sociability: females frequently affiliate with each other, and appeasement actions are quite common (Nishida and Hiraiwa-Hasegawa 1987). Although food sharing is more frequent in male-female pairs, it is not exceptional in female-female pairs as well, involving even non-related adult females (Hohmann and Fruth 1993); such cases would be almost impossible in chimpanzee communities (Kuroda 1984).
Egalitarian vs. Non-Egalitarian Primates: A Synthesis
On the basis of the database above a correlation matrix was created for eleven species. The reason we chose these species from the total list of 19 is to provide more equal representation of different primate genera: we chose four macaques species (two egalitarian: M. tonkeana and M. arctoides and two despotic: M. mulatta and M. fuscata); three species from genus Papio (P. anubis, P. papio, P. ursinus); Cercopithecus aethiops; and three great ape species (see Table 1). Needless to say that there is no way to reproduce a comprehensible 40×40 correlation matrix in a journal paper format. Hence, we decided to reproduce just a part of it. The results of statistical tests of correlation (Spearman's Rho and respective levels of significance) between two behavioral patterns (‘Demonstration of Submission’ and ‘Permissiveness of Non-subordinate Behavior towards Dominants’) and the rest of them (see Table 2) are presented below. We regarded the first variable as a clear indicator of an overall non-egalitarian pattern, whereas the second could be considered as a clear indicator of an overall egalitarian one. Hence, we assumed that the traits displaying a significant positive correlation with the first variable could be regarded as characteristics of the non-egalitarian organization, whereas the traits displaying a significant positive correlation with the second could be regarded as characteristics of the egalitarian one. Thus, the general behavioral ‘profile’ of egalitarian and non-egalitarian social organization irrespective of residence patterns and types of mating patterns was created (see Table 2).
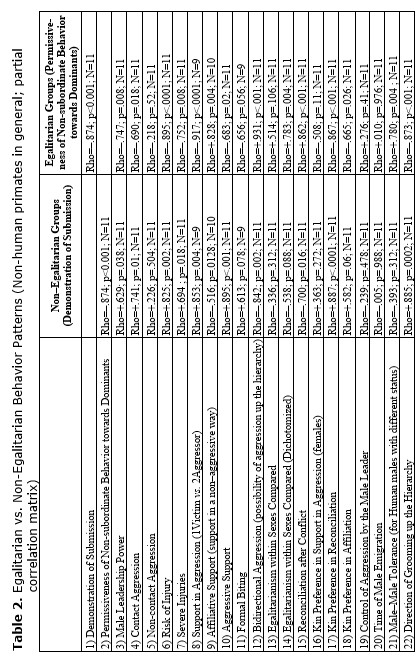
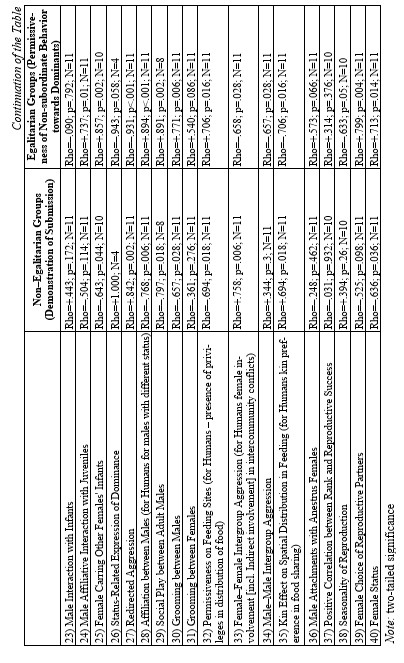
The results demonstrated that the following behavioral traits are highly correlated with egalitarianism and non-egalitarianism: male leadership power (i.e., if any group members ever try to attack or object the leader), contact aggression, risk of injury, severe injury, support in aggression (on the side of either victims or aggressor), affiliative support, aggressive support, formal biting, bidirectional aggression, egalitarianism within sexes compared (if the level of egalitarianism is the same for males and females), reconciliation after conflict, kin preferences in reconciliation, kin preferences in affiliation, direction of grooming up the hierarchy, female carrying other females' infants (i.e., permissiveness in access to infants for other females, non-kin included), redirected aggression, social play between males, grooming between males, permissiveness on feeding sites, female-female intergroup aggression, male-male intergroup aggression, kin effect on spatial distribution in feeding, female choice of reproductive partner and female status (see Table 2). In all cases the directions of ties were alternative in case of egalitarianism compared to non-egalitarianism. Some traits in our sample bear correlations for egalitarian species only: male-male tolerance, male affiliative interaction with juveniles, grooming between females, male attachments with anestrus females (positively), and seasonality of reproduction (negatively) (see Table 2). No significant correlation was found between the type of dominance style and the level of non-contact aggression, control of aggression by the male leader, time of male emigration, male interaction with infants and positive correlation between rank and reproductive success (see Table 2). It is necessary to admit that the absence of correlation in the case of the ‘time of male emigration’ is a consequence of a high proportion of male phylopatric species in the sample. If the whole sample of 19 species is under analysis, correlations are high and in predicted directions (for non-egalitarian groups: Rho = –0.466; p < 0.022; for egalitarian groups: Rho = +0.391; p < 0.049). It means that in non-egalitarian species males usually emigrate earlier than in egalitarian ones.
Non-egalitarianism
vs. Egalitarianism Among
Non-Human Primates and Simple
Hunter-Gatherers Compared
Is it possible to detect a similar set of behavioral patterns among humans? Let us start with the simple nomadic hunter-gatherers. As is known, Woodburn and Artemova use the Australian Aborigines as archetypical example of non-egalitarian hunter-gatherers, whereas one of the typical examples of the egalitarian hunter-gatherers is the Bushmen (Woodburn 1980, 1988a, 1988b; Artemova 1987, 1993, 2000). Let us study now how much the behavioral patterns of the Aborigines are similar to the ones of the non-egalitarian primates, and how much the behavioral patterns of the Bushmen are similar to the ones of the egalitarian primates.
1a. As was mentioned above, the non-egalitarian primate groups are characterized by the developed patterns of the demonstration of submission. The Australians showed a marked system of the demonstrations of submission, for example during the initiations. All Australian aboriginal tribes had elaborate, prolonged and painful initiation rites during which the adolescents explicitly and in highly ritualized manner submitted themselves to tooth evulsion, septum (nose) – piercing, circumcision, sub-incision, fire ordeals, scarification, fingernail pulling (about Aranda see Strehlow 1947: 112–4), beatings, blood-letting, pulling out of hair, hunger ordeals (boys) and to ritual defloration (and subsequent multiple sex relations), septum piercing, genital circumcision (girls), removal of finger joints, scarification (Myers 1980: 207; Piddington 1932–1933: 46–87; Engelhart 1998: 58–66; Tindale 1935–1936: 200–224; Berndt and Berndt 1964: 136–157; Layton 1986: 41–45). All these forms of behavior may be classified – on the part of the initiated – as demonstrations of submission (see especially Spencer and Gillen 1927: 457–465; Berndt and Berndt 1964: 144; Keen 1994). Initiations took form of stages each lasting up to several months and joined into chains which extended through years, for example, ‘The people at Ooldea had their novices spent nine to twelve months in seclusion…’ (Berndt 1942–1943: 262; see also Berndt and Berndt 1964: 136–157; Engelhart 1998: 58–66).
1b. The egalitarian primate groups are characterized by the absence of such patterns. The !Kung Bushmen had very short in duration (about two weeks) and loosely organized initiations for boys (/choma) which did not include any enduring and painful trials or painful surgical operations apart from making small vertical cut between eyebrows (or on a chest) and rubbing the cut with some ash powder (Bjerre 1963: 122–127; Marshall 1976: 130–131). There are grounds to suppose that the /choma among the !Kung diffused from the Bantu at the beginning of the 20th century (Idem. 1965: 264). The Southern Bushmen (/Xam and ¹Khomani) did not have initiation for boys (Barnard 1992: 80–81; Hewitt 1986: 279–286).[1] In general, in contrast with the Australian Aborigines, the Bushmen do not appear to have had any developed patterns of the demonstration of submission.
2a. The non-egalitarian primate groups are characterized by lack of permissiveness of non-subordinate behavior towards dominants. Among the Australian Aborigines there were developed systems of punishment for non-subordinate behavior towards dominants. Wives were being physically punished by the husbands for almost any attempt of direct contradiction (e.g., Berndt and Berndt 1964, 1970; White 1978). In addition to that the seniors could decide to punish (often by death) various sorts of culprits under the pretext of the breaching the norms of sacral behavior. Clear enough this served as a powerful means of securing subordinance, since the breach of the sacred law might serve as only a pretext under which various real causes of the senior dissatisfaction could lie. The senior men then could choose among the younger men a special agent for the execution of a punishment (including even a death penalty [Berndt and Berndt 1964: 286–289; Spencer and Gillen 1927]).
2b. The egalitarian primate groups are characterized by relative permissiveness of non-subordinate behavior towards dominants. Such behavior was quite possible among the Bushmen, for example, a wife could threaten to leave a husband if he took a fancy to get a second wife. ‘They just said they did not want to be second wives and they told of protests and quarrels that attended the husbands' proposals to take second wives’ (Marshall 1976: 265). We may give the following example of non-subordinate behavior (which also illustrates bidirectional aggression):
On one occasion, Gao Beard coerced his young second wife into going with him when she wanted to stay visiting her parents. He coerced her swiftly and decisively by snatching her baby from her arms and walking off with him. In a flash, the wife ran a few steps and hit him on the head with her digging stick, then she went around in a circle, stamping her feet in great high stamps like an enraged samurai in a Japanese print, then she followed her husband (Ibid.: 312).
One should emphasize here that the husband did not react to the blow.
3a. The non-egalitarian primate groups are characterized by the high level of the male leadership power. The system of male-dominated social ranking was ingrained in the social pattern of all aboriginal groups. For the Western Arnhem Land tribes (but the traits demonstrated below existed throughout Australia) the basic traits of this ranking were outlined by R. and C. Berndts as follows:
The basis of social control was acceptance of the normative order… The essence of the system was assent, agreement on the rightness of the rules and unquestioning acceptance of the supernatural or spiritual order, which supported them. The key to its practical working out lies in the social control and organization of sacred ritual – that is, (i) domination by senior men, (ii) younger to middle-aged men acting as junior executives and handling most of the active work, (iii) the women, also differentiated with respect to the age and experience, having limited authority and limited autonomy in certain fields, and (iv) responsibility for pre-adolescent children resting with adults of either both sexes according to circumstances. This agreement, tightly organized in the ritual context, was superimposed more loosely on the entire range of activities outside it (Berndt and Berndt 1970: 150–151).[2]
3b. The egalitarian primate groups are characterized by low level of the male leadership power.
Among the !Kung Bushmen the male leadership was manifested rather moderately, whereas the power of the community formal male leader (k''hau n!a) was virtually non-existent. Communities (bands) comprised quite autonomous extended and even nuclear families outside which power decisions were exercised only by way of communal consensus (Marshall 1976: 191–195; Thomas 1959: 183). Similar systems of power were characteristic of the other Kalahari Bushmen (Silberbauer 1981: 28–34; Barnard 1992: 108–109).
4–6a. The non-egalitarian primates are characterized by high levels of contact aggression, risks of injury, and severe injuries. There are direct comparable numerical data of the homicide levels for the following communities: the Vanindiljaugwa ‘tribe’ of pre-contact South-Eastern Australia (non-egalita-rian), Murngin ‘tribe’ of North-Eastern Arnhem Land (non-egalitarian) and the !Kung of the so-called Nyae Nyae region in the adjacent parts of north-eastern Botswana and north-eastern Namibia (egalitarian).
For the Wanindiljaugwa (Victoria State, Australia) the calculations of Blainey are as follows: the size of the tribe is 280–420 persons; average level of the casualties in internal and external warfare: one person to 270 persons for the period of one year (Blainey 1975: 109–111). The calculations of Blainey are based on the published recollections of the runaway convict William Buckley who lived among Wanindiljaugwa in 1804–1835 (Blainey 1975: 106; Buckley 1852: 14, 85) in totally pre-contact situation. Reliability of Buckley's data is corroborated by the data of the missionary L. Threlkeld (1974). The book of the latter was published in 115 years after the time of his observations (1824–1859). A total list of casualties mentioned by W. Buckley comprises minimum of 50 persons for 30 years including ten women and twelve children.
For the Murngin people (North-Eastern Arnhem Land; data of W. L. Warner [1937: 158–163; Blainey 1975: 109–111): period of observation (data from informants): 1909–1929; size of the population: c. 3000 persons; total casualties: 200 persons; homicide level: 1/300 for one year, that is, approximately the same as conservative calculations for Wanindiljaugwa.
Other ecologically productive regions of aboriginal Australia also had high
levels of contact aggression (Berndt R. and Berndt C. 1964: 299–302). R. and
C. Berndts remark that ‘the most highly organized warfare in Aboriginal Australia
is found in north-eastern and north-central Arnhem Land’ (Ibid.). One should
note here that these regions differed, at the time of observations, from Western
Arnhem Land mostly in lesser degree of acculturation which in the given historical
context lowered the levels of native homicide. Still these levels were, among the
Murngin of north-eastern Arnhem Land slightly lower than in south-eastern Australia
among the absolutely non-acculturated Wanindiljaugwa of 1804–1835. It shows that
most probably the levels of contact aggression in pre-contact Australia were even
higher than the ones generally ethnographically reconstructed.
4–6b. The egalitarian primates are characterized by low levels of contact aggression, risks of injury, and severe injuries. In the Western Desert of Australia (whose groups in contrast with the rest of Aboriginal Australia were characterized by more or less egalitarian social organization [e.g., Yengoyan 1970: 81–85; 1976: 126–128; Liberman 1985: 10–15, 27–31, 101–104]) levels of aggression were significantly lower than the ‘average’ Australian ones (Berndt 1965: 200; Tindale 1972: 104; Gould 1969: 149; Tonkinson 1974: 18; Terry 1928: 24).
For the !Kung Bushmen of the Nyae Nyae and ‘Dobe’ (the latter region is called ‘/Kangwa’ in Botswana), we have the following data of R. Lee (1979: 383): period of observation (data from informants): 1920–1955; size of the population: c. 560 persons; total casualties: nine persons for 30 years; homicide level: 0,15/250 for one year. Conclusion: homicide level among the !Kung is more than six times lower than in pre-contact Victoria or north-east Arnhem Land. Non-numerical data on low levels of homicide exist, as well, on the other Kalahari Bushmen groups (G/wi, Nharo, !Ko: Silberbauer 1972: 303–305; 1981: 192–193; Guenther 1986: 7, 40–45; Barnard 1992: 134–141). The !Kung norms of social behavior are strongly oriented towards avoiding and disapproving any sort of conflict: ‘The !Kung' s dread of fighting is absolutely overt and deliberate, they speak about it often and each manifestation of disagreement in the band makes them feel uneasy. … their tendency to avoid animosity and disagreement makes them follow the unspoken conducts of behavior very strictly’ (Marshall 1976: 311, see also pp. 53, 188, 200, 282, 312; Lee 1972: 359–360). In north-east Arnhem Land the corresponding socialization norms are aimed at producing a belligerent adult male (Berndt 1964: 289; Chaseling 1957: 87–88; Lamilami 1974: 8–10, 73–74, 87; for the Tiwi see Hiatt 1959: 188–191).
7–8a. The non-egalitarian primates are characterized by low level of affiliative support and high level of aggressive support. Among the Australian aborigines the situation is much more complicated: fights between women often result (at least among the Gunwinggu of Western Arnhem Land) from the efforts of the mothers to protect their small sons and daughters in fights. A quarrel between children (quite a common thing) thus may naturally develop into a duel between the two mothers, armed with digging sticks, with a group of the onlookers in a circle and adult men intervening only when risk of serious injuries arises (Berndt R. and Bernd C. 1970: 166–167; Berndt R. and Berndt C. 1964: 135).
The fights between adult males (and females) of the same community quite commonly took form of formal duels with the onlookers. Such duels (between adult males and formal fights between boys) were witnessed even for Ayers Rock Yankuntjantjara who were much less aggressive than Arnhem Land Aborigines (Porteus 1931: 168–169). Intervention of the adult males only in situations when risk of serious injuries infliction arose may well be qualified as lack of affiliative support in comparison with the rigid suppression of all (even potential) fighting among the Bushmen.
7–8b. The egalitarian primates are characterized by high level of affiliative support and low level of aggressive support. Among the !Kung actually observed by Lorna Marshall and John Marshall all potentially dangerous conflicts were resolved by means of ‘talking’. For the period of 17,5 months of continuous observations (1952–1953) with two bands and visitors (up to 75 people altogether) Lorna Marshall witnessed only four serious flare-ups and also heard about three more in the neighboring bands. All these conflicts were resolved before they took serious course (a murder is technically an easy thing for a Bushman hunter: poisoned arrows are always at hand). The policy of deliberate quieting of any quarrel may be regarded as a form of affiliative support of the victim of aggression. All authoritative and present-minded persons might take part in this process. A typical example may look like follows: John Marshall reports of a case when two half-sisters of an enraged hunter ready to snatch his bow and arrows ‘threw themselves on him, pulled him away and sat down beside him holding his arms. He quieted down…’ (Marshall 1976: 282). Another example: ‘A husband and a young widower had almost fought over the young widower's attentions to his wife. The fight had been averted by the husband's running away’ (Ibid.: 281). The conflict was later settled by a leader (¹Toma) of the other band. The tendency to avoid fighting as much as possible gives at least indirect evidence of the rarity of aggressive support.
9. Bidirectional aggression (low level in non-egalitarian and high level in egalitarian primates). As was mentioned above, the aggression against dominants on the part of subordinates was most severely punished among the non-egalitarian Australian aborigines in contrast with the social life of the Bushmen. A typical example of such type of aggression without any punishment among the !Kung was quoted above (2b. – Ibid.: 312). Another (extreme) example: ‘Two men at /Gam quarreled about their plans and arrangements to make a gathering trip. One was Tsamgao, the father of ¹Toma. The young son of the other man became frightened for his father and wanted to protect him. He took a poisoned arrow from his father's quiver and shot and killed Tsamgao’ (Ibid.: 190). Again, no retaliation was made, though ¹Toma had eventually to leave the band (he joined into another band, about 200 km to the north).
10. The non-egalitarian primates are characterized by low level of male–male tolerance. The egalitarian primates are characterized by high level of male–male tolerance. The non-egalitarian Australians are characterized by low level of male–male tolerance (for males with different status). The egalitarian Bushmen are characterized by high level of male–male tolerance (for males with different status), see # # 1, 4 and 5b above.
11. Male affiliative interactions with juveniles (low level for non-egalitarian and high level for egalitarian primates). In the non-egalitarian Aboriginal societies, senior males who dominated social life (see No 1) or fully initiated males almost certainly were not likely to participate in games with children, although there is a lack of direct data in this respect. However, in several thick general monographs we could not find a single mentioning of the initiated male adults playing games (e.g., Berndt and Berndt 1964, 1970; Spencer and Gillen 1927, 1913; Elkin 1954).
This trait is explicitly mentioned for the egalitarian !Kung Bushmen: ‘Adults played with the children and teenagers when they wished. Teenagers played with younger children’ (Marshall 1976: 314; 1965: 264). Lorna Marshall gives data on several concrete games played both by boys and young men (Marshall 1976: 332–337; 345, 351). In one case stick throwing is mentioned as a favourite pastime among Nharo of not only boys and young men but also of the middle-aged men (Marshall 1976: 337; Bleek 1928: 21).
12. Female carrying other females' infants is more typical for egalitarian than non-egalitarian primates. This trait is much more typical for the egalitarian Bushmen than the non-egalitarian Australians. Among the !Kung there are no preferential female categories in this respect. All females carry other's babies. When babies ‘are not in their mother's arms or tied to their sides, they are in someone else's arms… The !Kung never seem to get tired of their babies. They dandle them, kiss them, dance with them and sing to them. The older children make playthings of the babies. The girls carry them around… The boys also carry the babies around, give them rides and drag them on carosses [a favourite game]’ (Marshall 1976: 315–318). Among Aborigines (in Western Arnhem Land) certain classes of female relatives are singled out. Apart from mother's mother these are her sister, classificatory sisters of the mother and co-wives (Berndt and Berndt 1970: 158; 1964: 129).
13. Affiliation between males (for males with different status) is more typical for egalitarian than non-egalitarian primates. This trait is obviously more marked among Bushmen since they lack in comparison with Australian Aborigines, sharp male (and female, incidentally) status distinction (see No 1).
14. Social play between adult males is much more typical for egalitarian than non-egalitarian primates. Social play between adult males is much more typical for the egalitarian Bushmen than the non-egalitarian Australians, see # 8 above.
15. The non-egalitarian primates are characterized by low level of permissiveness on feeding sites. The egalitarian primates are characterized by high level of permissiveness on feeding sites. Among Australian Aborigines ‘The Gunwinggu boys were told in the opening session of their initiation not to eat certain foods such as barramundi, catfish, goanna, tortoise, crocodile, bandicoot and many others. They were also forbidden to eat young or fat kangaroo...’ (Berndt R. and Berndt C. 1970: 131, 163, 52, 115). The women also could be subject to food restrictions during initiations (Ibid.: 132). All throughout Aboriginal Australia there were food restrictions for the men with non-full ritual statuses and women (Ibid.: 134; Engelhart 1998: 81–84; Berndt 1964: 125, 150, 153–154, 156, 203, 220, 241; Meggitt 1962: 283; Spencer and Gillen 1899: 256, etc.). In Western Arnhem Land ‘Older men directly or indirectly decided who had access to foods through ritual restrictions which they could impose or waive, declaring them dhuyu (“sacred, taboo”)’ (Berndt R. and Berndt C. 1970: 98). There is no data on food restrictions for Bushmen boys while being initiated but in view of the short duration of the procedure (in comparison with Australia, see Variable 1) there are no grounds to doubt the existence of a sharp distinction between Australian Aborigines and Bushmen in the analyzed parameter.
Among the Bushmen no food was secured specially for men or men with the high status. Although at least one case of food taboo is mentioned by L. Marshall, ostrich eggs must not be eaten ‘by men and women from the age of puberty till they are old enough to have five children…children and older people may eat them’ (Marshall 1976: 98) – we can see that there is no real status discrimination in this taboo.
16. Female–female inter-group a ggression is more typical for non-egalitarian rather than egalitarian primates. Aboriginal women might occasionally fight alongside with men (Wanindiljaugwa: Buckley 1852: 36–37) or take part in a stealth vengeance raid (Aranda: Spencer and Gillen 1927, v. 2: 461–464). In addition, in some Australian tribes women ritually imitated aggression taking up spears when the men would come to ‘grab’ boys for the initiations (Engelhart 1998: 101; Warner 1937: 251; Piddington 1932–1933: 70). The existence of armed duels between women may also serve as indirect evidence to the supposition that in desperate situations they could give military aid to their men.
Bushmen women (direct data for !Kung and Nharo) were fleeing at such conflicts (Baines 1864: 137, 334–335, 358).
17. The non-egalitarian primates are characterized by high level of male–male inter-group aggression. The egalitarian primates are characterized by low level of male–male inter-group aggression. The non-egalitarian Australians are characterized by high level of male–male inter-group aggression. The egalitarian Bushmen are characterized by low level of male–male inter-group aggression, see # 3 above.
18. The egalitarian primates are characterized by a higher level of female choice of reproductive partners than the non-egalitarian primates. In both Australians and Bushmen there is a practice of early marriages, in which the decision of betrothal is made by parents. Among !Kung, however, bride's voice matters equally in contrast with Australia (compare, e.g., Whyte 1978 and Marshall 1976: 266).
19. The non-egalitarian primates are characterized by male dominance over females and the egalitarian primates – by its virtual absence. This trait was highly developed in the traditional Aboriginal social pattern (for which there is abundant evidence, see e.g., Berndt R. and Berndt C. 1970: 110, 150–151, 186, 232, 235; Whyte 1978; Artemova 1987) and is obvious from almost all the analyses made above (see, e.g. variable No 1). It is virtually absent among the Bushmen.
Thus, out of 30 variables characterized by significant correlations with the non-egalitarian/egalitarian social organization in non-human primates, the ‘Australians–Bushmen’ comparison showed direct and distinct parallelism in 19 cases. At present five variables are too difficult to evaluate with the necessary degree of certainty but they are quite likely to join the above described parallelism. These are: ‘Kin Preference in Support in Aggression’, ‘Kin Preference in Reconciliation’, ‘Kin Preference in Reconciliation’, ‘Kin Effect on Spatial Distribution in Feeding (for Humans kin preference in food sharing’) (in view of the possible less close consanguine relatedness of the members of the bands among Bushmen), ‘Redirected aggression’ and ‘Male Attachment with Anestrus Females (~ friendly sex-free relations between men and women)’. Four variables are obviously irrelevant for humans (the ones related to grooming and ‘formal biting’).
There is only one variable (out of 30!) which does not show any significant difference between the Australians and the Bushmen: ‘Reconciliation after Conflict’ (Berndt 1965: 200; Tindale 1972: 104; Gould 1969: 149; Tonkinson 1974: 18; Terry 1928: 24; Lee 1979: 383; Silberbauer 1972 303–305; 1981: 192–193; Guenther 1986: 7, 40–45; Barnard 1992: 134–141). And there are no variables at all showing a significant difference in the direction opposite to the predicted one.
Non-Egalitarianism vs.
Egalitarianism Among
Non-Human Primates and Humans Compared
Finally, let us compare the patterns of egalitarianism/non-egalitarianism among the non-human primates on the one hand, and the humans in general on the other. In order to do this, we decided to compare the correlation matrices of the egalitarianism/non-egalitarianism characteristics for both groups of social systems. For the humans we opted to use the published coded data for the Standard Cross-Cultural Sample (SCCS: Murdock and White 1969; SCCS 1999; Divale et al. 2002). Of course, the data for most variables mentioned above are not available for the SCCS cultures. Hence, we had to restrict ourselves to those variables for which those data were available. These variables are: ‘Permissiveness of Non-subordinate Behavior towards Dominants’, ‘Bidirectional Aggression’, ‘Male Leadership Power’, ‘Contact Aggression’, ‘Kin Preference in Affiliation’, ‘Female Status’, ‘Female Choice of Reproductive Partners’. Three out of seven chosen variables (bidirectional aggression, contact aggression and kin preference in affiliation) are widely used as indicators of ‘despotism’ and ‘egalitarianism’ in non-human primates (Butovskaya 1993; Matsumura 1999); the rest four were chosen because they are highly correlated in the prediction directions with these three, as well as because for all seven parameters it was possible to find analogues for human societies.
For the non-human primates the correlation matrix for those variables is as follows:
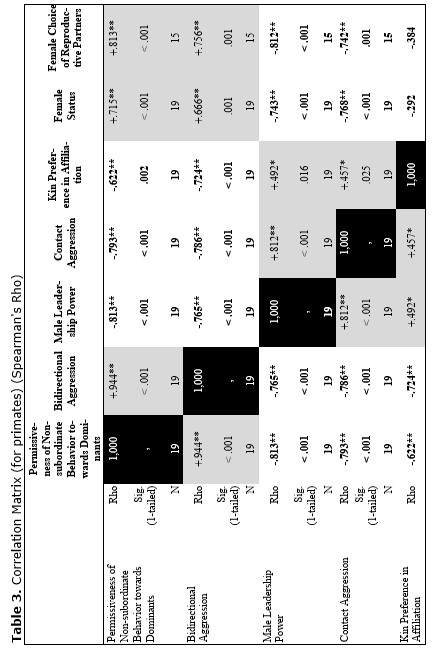
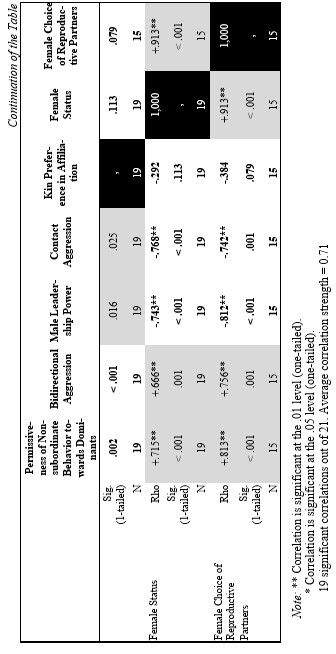
Now compare this correlation matrix with the respective correlation matrix for the humans. But before we reproduce this matrix, it is necessary to make a few comments.
First of all, one should emphasize that for none of the variables analyzed above we have complete correspondences in the SCCS database. However, it seems possible to find rather close analogues to them. Hence, we decided to use the following set of analogues.
1) ‘Permissiveness of Non-subordinate Behavior towards Dominants’ ~ ‘Political Participation: Extensivity of Adult Participation’ (Ross 1983, 1986; SCCS Variable 766). It is not immediately clear that the second variable corresponds closely to the first; however, it does become clear if one looks at how this variable was coded:
1 = Widespread: decision making forums open to all adults and involvement is relatively great;
2 = High or some: widespread for some groups but exclusion of others on the basis of gender, age or kinship status;
3 = Moderate: some consultation is present but low input from community;
4 = Low or non-existent: Leaders make most decisions and involvement of average person is highly limited or absent.
We re-coded this variable as follows:
4 = Widespread: decision making forums open to all adults and involvement is relatively great;
3 = High or some: widespread for some groups but exclusion of others on the basis of gender, age or kinship status;
2 = Moderate: some consultation is present but low input from community;
1 = Low or non-existent: Leaders make most decisions and involvement of average person is highly limited or absent.
It is easy to see that the situation implied by the maximum value of the re-coded variable corresponds to the situation when the non-subordinate behavior towards dominants is possible by the definition, whereas it becomes less and less possible with every lower value.
2) ‘Bidirectional Aggression’ ~ ‘Checks on Leaders' Power’ (Ross 1983, 1986; SCCS 1999: Variable 761). We believe that the first variable corresponds to the second one as the presence of the possibility of the aggression up the hierarchy is tantamount to the presence of the checks on leaders' power, whereas any effective checks on the leaders' power implies the possibility of (normally non-contact) aggression towards them on the part of their subordinates; for example the possibility to dispute their decision, to reelect them, etc.
3) ‘Male Leadership Power’ ~ ‘Leaders' Exercise of Authority’ (Ross 1983, 1986; SCCS 1999: Variable 763).
4) ‘Contact Aggression’ ~ ‘Conflict [Social or Political] in the Local Community’ (Ross 1983, 1986; SCCS 1999: Variable 767). It is not immediately clear that the second variable closely corresponds to the first; however, it does become clear if one looks at how this variable was coded:
1 = Endemic: a reality of daily existence (e.g., physical violence, feuding, bitter factionalism);
2 = High: Conflict is present but not a pervasive aspect of daily life;
3 = Moderate: Disagreements and differences do not result in high violence or severe disruption;
4 = Mild or rare.
We re-coded this variable as follows:
4 = Endemic: a reality of daily existence (e.g., physical violence, feuding, bitter factionalism);
3 = High: Conflict is present but not a pervasive aspect of daily life;
2 = Moderate: Disagreements and differences do not result in high violence or severe disruption;
1 = Mild or rare.
It is easy to see that the situation implied by the maximum value of the re-coded variable corresponds to the situation when the contact aggression is most frequent, whereas it becomes less and less frequent with every lower value.
5) ‘Kin Preference in Affiliation’ ~ ‘Corporate Kin Groups’; coded on the basis of the Ethnographic Atlas database (Murdock 1967, 1981; Murdock et al. 1986, 1990, 1999).
6) ‘Female Status’ ~ ‘Ownership or Control of the Use of Dwellings’ (Whyte 1978, 1985: stds23.dat; SCCS 1999: stds23.sav). We chose this variable from the list of 60 characteristics of the female status, the coded data for which had been published by Whyte, as, according to our earlier experience, this variable had turned out to be the most reliable.
7) ‘Female Choice of Reproductive Partners’ ~ ‘Romantic Love as a Basis of Marriage’ (Rosenblatt 1967, where ratings for 75 SCCS cultures are published). Here we used a dichotomized version of the variable (de Munck and Korotayev 1999; for the dichotomization justification see pp. 268–269; for the dichotomized ratings see pp. 274–276). We assumed that among the humans the romantic love as a basis of marriage would strongly correlate with the female choice of reproductive partners.
We also made additional codings for some SCCS variables for the following SCCS cultures: Aranda (on the basis of Spencer and Gillen 1899; Strehlow 1947); Armenians (Tolstov 1962); Babylonians (Diakonoff 1983); Balinese (Geertz and Geertz 1975); Botocudo (Ehrenreich 1887; Nimuendajú 1946); the Chinese (Yang 1945); Egyptians (Ammar 1954); Hadza (Bleek 1931; Woodburn 1968a, 1968b); the Japanese (Embree 1939; Norbeck 1954); Koreans (Lee 1975); !Kung (Bjerre 1960; Marshall 1976; Thomas 1959; Wilhelm 1954); Mbuti (Gusinde 1956; Schebesta 1933, 1948; Turnbull 1961, 1965a, 1965b); Russians (personal observations of the authors); Tiwi (Basedow 1913; Goodale 1962, 1971; Hart 1930; Mountford 1958); the Turks (Makal 1954); Uttar Pradesh (Gore 1965); West Punjabi (Zekiye 1957). As the modern egalitarian western societies are not represented in the SCCS at all, we decided to make additional coding for three of them on the basis of our personal observations (the English: the Withington neighborhood in Southern Manchester, observations of 1991–1994; the Dutch: the community of Utrecht, observations of 1994–1996; Swiss: Zürich, observations of 1991–1993). The total dataset used to produce the correlation matrices below is reproduced in the Appendix.
The correlation matrix for the whole world-wide cross-cultural sample looks as follows (see Table 4).
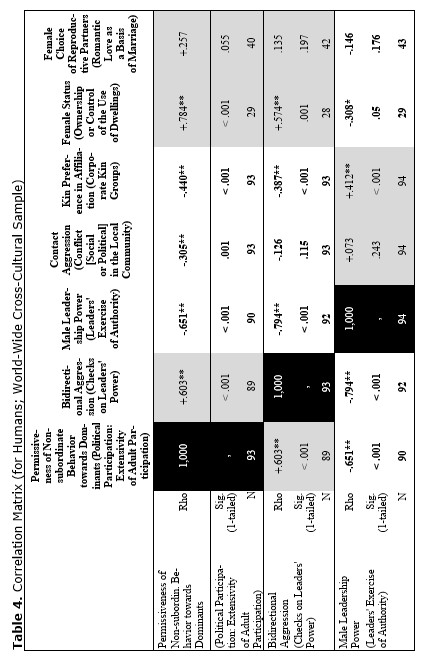
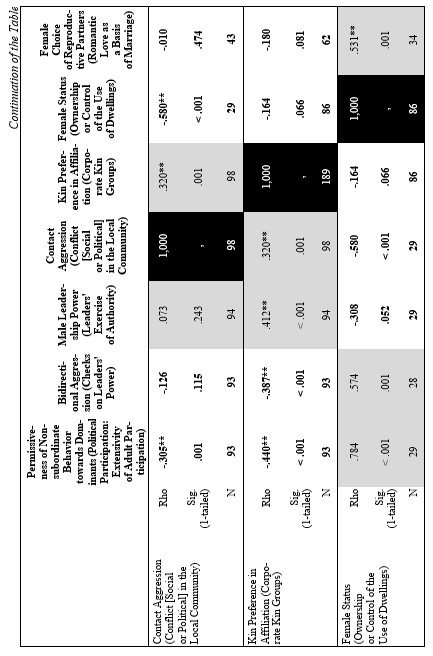

Yet, the analysis of the egalitarian vs. non-egalitarian behavioral patterns among the Bushmen vs. Australians would suggest that those correlations must be stronger among the simple nomadic hunter-gatherers. And, indeed, this is the case (see Table 5). For the measurement of cultural complexity we used the cultural complexity index (CCI) elaborated by Murdock and Provost (1971, 1985; SCCS 1999: Variables 149–158, where the CCI values for all 186 SCCS cultures are published).
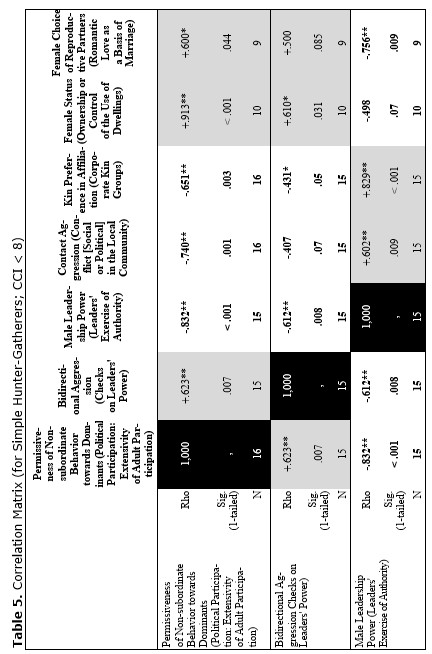

As we can see, the correlation matrix for the simple nomadic hunter-gatherers turn out to be quite close to the one for the primates. This is not really surprising, as they live in conditions which are most close to the one within which the human biogram was formed. Hence, one would expect that the behavioral predispositions among them would affect the actual behavior more significantly than in more complex societies where people would live in the conditions radically different from the ones within which the human biogram was formed.
Now let us analyze the correlation matrices for the societies with medium and high levels of cultural complexity. But, firstly, we have to answer some questions. Indeed, one may wonder if the egalitarianism vs. non-egalitarianism dichotomy is relevant for the medium and high complexity cultures. Do not egalitarian societies disappear altogether as soon as the medium level of cultural complexity is reached? Does not the egalitarian social system occur among the simple cultures only?
Indeed, some of the most popular unilineal socio-evolutionist schemes make us believe in it. Of course, one would have to admit that the egalitarian societies already disappear at the medium levels of cultural complexity if one defines them as ‘societies in which all persons of a given age-sex category have equal access to economic resources, power and prestige’. However, even if we define the egalitarian societies as those characterized by ‘weakly linear or shallow dominance relations’, the standard unilinear evolutionist schemes would still imply the same.
Let us take, for example, the famous Sahlins/Service staircase of the ‘levels of cultural integration’ (Service 1962[1971]; its outline is, however, already contained in Sahlins 1960: 37): band – tribe – chiefdom – state (this scheme is till now reproduced in the majority of the most widely used textbooks in cultural anthropology [Ember and Ember 1999: 375–397; Haviland 1991: 530–539; Kottak 2000: 356–380, etc.]).
The scheme implies precisely the above-mentioned interpretation whereas less hierarchized societies are automatically considered to be less developed than more hierarchical ones. It implies that the growth of cultural complexity (at least up to the stage of the agrarian state) is inevitably accompanied by the growth of inequality, stratification, the social distance between the rulers and the ruled, the ‘authoritarianism’ and hierarchization of the political system, decrease of the political participation of the main mass of population, etc. This is just due to the constant growth of the relative importance of highly lineal and rigid dominance relations.
In general, the transition from the egalitarian stage of social evolution to the ‘non-egalitarian’ one seems to be identical with the transition from the supposedly universal stage of the ‘tribe’ to the supposedly universal stage of the ‘chiefdom’. Thus, compare Service's descriptions of the ideal models of the tribe and the chiefdom which are represented by him as universal stages of sociopolitical evolution. How does Service define the main differences between the political organization of the tribe and the chiefdom? The political organization of the tribe is described in the following way:
Leadership is personal ... and for special purposes only in tribal society; there are no political offices containing real power and a `chief' is merely a man of influence, a sort of adviser. The means of tribal consolidation for collective action are therefore not governmental... Tribe ... is composed of economically self‑sufficient residential groups which because of the absence of higher authority take unto themselves the private right to protect themselves. Wrongs to individuals are punished by the corporate group, the `legal person'. Disputes in tribal society tend to generate feuds between groups[3] (Service 1962 [1971]: 103).
The socio-political organization of the chiefdom is characterized by Service as follows:
‘The great change at the chiefdom level is that specialization and redistribution are no longer merely adjunctive to a few particular endeavors, but continuously characterize a large part of the activity of the society. Chiefdoms are redistributional societies with a permanent central agency of coordination’ (Service 1962 [1971]: 134).
When chieftainship becomes a permanent office in the structure of society social inequality becomes characteristic of the society, followed finally by inequality in consumption... The creation of the hereditary office of chief, with its high status for the person who occupies it, naturally carries the possibility of other statuses of high degree... A chief's high status raises the status of every member of his family above ordinary families, and ultimately that of the families in his local kin group to some extent... A chief necessarily has a ‘nobility’, even though they are only his own family... A further important feature lies in the chief's ability to plan, organize and deploy public labor (Ibid.: 139–40).
‘A chiefdom is in a sense pyramidal or cone‑shaped in structure...’ (Ibid.: 142).[4]
Finally, Service describes some important differences between the chiefdom and the tribe:
‘...A chiefdom differs radically from a tribe or band not only in economic and political organization but in the matter of social rank –... tribes are egalitarian, chiefdoms are profoundly inegalitarian’ (Ibid.).
The most distinctive characteristic of chiefdoms as compared to tribes ... is ... the pervasive inequality of persons and groups in the society. It begins with the status of chief as he functions in the system of redistribution. Persons are then ranked above others according to their genealogical nearness to him. Concepts involving prescriptions, proscriptions, sumptuary laws, marriage rules and customs, genealogical conceptions, and etiquette in general combine to create and perpetuate this sociopolitical ordering, and in turn have an effect on social structure and status terminology and etiquette behavior. A charismatic ephemeral leader of the type found in tribes ... has the functions and attributes that result from his own capabilities.[5] An ‘office’, on the other hand, is a position in a sociopolitical structure that has ascribed functions and conventional attributes no matter who occupies it, etc. (Ibid.: 145–146).
Of course, for the medium-range societies it seems possible to find a negative correlation between the egalitarianism and the cultural complexity. But, no doubt, this is just a correlation and by no means a functional dependence. Needless to say, this correlation implies a perfectly possible line of socio-political evolution – from an egalitarian, acephalous band, through a big-man village community with much more pronounced inequality and political hierarchy to an ‘authoritarian’ village community with a strong power of its chief (found, e.g., among some Indians of the North-West Coast, see, e.g., Carneiro 2000), and then through the ‘true’ chiefdoms having even more pronounced stratification and concentration of the political power in the hands of the chief, to the complex chiefdoms where the political inequality parameters reach a qualitatively higher levels and finally to the agrarian state where all such parameters reach their culmination (though one could move even further, up to the level of the ‘empire’ [e.g. Adams 1975]). However, it is very important to emphasize that at each level of the growing political and cultural complexity one could easily find evident alternatives to this evolutionary line (see, e.g., Artemova 2000, 2003; Bondarenko et al. 2002, 2004, 2011; Bondarenko 2007, 2008, 2014, 2020; Crumley 2007; Grinin and Korotayev 2009, 2011, 2017; Korotayev 2000, 2006, 2020; Kradin 2006, 2008, 2011a, 2011b, 2014, 2018, 2019, 2020; Small 2019, 2020).
The egalitarian vs. non-egalitarian alternatives of social evolution among simple nomadic hunter-gatherers were discussed above.
At the next level of the political complexity we can also find communities with both hierarchical and non-hierarchical political organization. One can mention, for example, the well-known contrast between the Indians of the Californian North-West and South-East:
The Californian chiefs were in the center of economic life, they exercised their control over the production, distribution and exchange of the social product, and their power and authority were based mainly on this. Gradually the power of the chiefs and elders acquired the hereditary character, it became a typical phenomenon for California... Only the tribes populating the North-West of California, notwithstanding their respectively developed and complex material culture, lacked the explicitly expressed social roles of the chiefs characteristic for the rest of California (Kabo 1986: 20).
One can also immediately recall the communities of the Ifugao (e.g., Barton 1919, 1922, 1938; Meshkov 1982: 183–197) lacking any pronounced authoritarian political leadership compared with the one of the communities of the North-West Cost but with a comparable level of overall socio-political complexity.
Hence, already at the levels of simple and middle range communities we observe several types of alternative socio-political forms, each of which should be denoted with a certain term. The possible alternatives to the chiefdom in the prehistoric South-West Asia, non-hierarchical (horizontally oriented) systems of complex acephalous communities with a pronounced autonomy of single family households have been analyzed recently by Berezkin who suggests reasonably Apa Tanis as their ethnographic parallel (Berezkin 1994, 1995a, 1995b, 1997, 2000; see also Grinin and Korotayev 2011, 2017). Frantsouzoff finds an even more developed example of such type of polities in ancient South Arabia in Wadi Hadramawt of the 1st millennium BC (Frantsouzoff 1995, 1997, 2000).
Another evident alternative to the chiefdom is constituted by the tribal organization. As is well known, the tribe has found itself on the brink of being evicted from the evolutionary models (Townsend 1985: 146; Carneiro 1987: 760). However, the political forms entirely identical with what was described by Service as the tribe could be actually found, for example, in medieval and modern Middle East (up to the present): these tribal systems normally comprise several communities and often have precisely the type of political leadership described by Service as typical for the tribe (Service 1971 [1962]: 103–104; Dresch 1984a: 39, 41; see also Chelhod 1970, 1979, 1985: 39–54; Dostal 1974, 1985, 1990: 47–58, 175–223; Obermeyer 1982; Dresch 1984b, 1989, 2016; Abu Ghanim 1985, 1990: 229–251; Weir 2007; Brandt 2017; etc.).
The point is that we are dealing here with some type of polity that could not be identified either with bands or with village communities (because such tribes normally comprise more than one community), or with chiefdoms (because they have an entirely different type of political leadership), or, naturally, with states. They could not be inserted easily either in the scheme somewhere between the village and the chiefdom. Indeed, as has been shown convincingly by Carneiro (see, e.g., 1970, 1981, 1987, 1991, 2000), chiefdoms normally arose as a result of the political centralization of a few communities without the stage of the tribe preceding this. On the other hand, a considerable amount of evidence could be produced suggesting that in the Middle East many tribes arose as a result of the political decentralization of the chiefdoms which preceded the tribes in time. It is also important to emphasize that this could not in any way be identified with a ‘regression’, ‘decline’ or ‘degeneration’, as we can observe in many of such cases that the political decentralization is accompanied by the increase (rather than decrease) of the overall social complexity (Korotayev 1995a, 1995c, 1995d, 1996a, 1996b, 1996c, 1997, 1998, 2000, 2003b, 2006, 2020). Hence, in many respects the tribal systems of the Middle Eastern type appear to be alternatives (rather than predecessors) of the chiefdoms.
We have argued elsewhere (Idem. 1995b) that in general there is an evident evolutionary alternative to the development of the rigid supra-communal political structures (chiefdom – complex chiefdom – state) constituted by the development of the internal communal structures together with the soft supra-communal systems not alienating the communal sovereignty (various confederations, amphictyonies, etc.). One of the most impressive results of the socio-political development along this evolutionary line is the Greek poleis (on the statelessness of this type of political systems see Berent 1994, 1996, 2004, 2006; see also Dozhdev 2004; Small 2019, 2020) some of which reached overall levels of complexity quite comparable not only with the ones of chiefdoms but also with the one of states.
The ‘tribal’ and ‘polis’ series seem to constitute separate evolutionary lines with some distinctive features: the ‘polis’ forms imply the power of the ‘magistrates’ elected in one or another way for fixed periods and controlled by the people in the absence of any formal bureaucracy. Within the tribal systems we observe the absence of any offices whose holders would be obeyed simply because they hold posts of a certain type and the order is sustained by elaborate mechanism of mediation and the search for consensus.
There is also a considerable number of other complex stateless relatively egalitarian polities (like the ones of the Cossacks of Ukraine and Southern Russia till the end of the 17th century [Chirkin 1955; Rozner 1970; Nikitin 1987; etc.] or the Icelandic polity of the ‘Age of Democracy’ till the middle of the 13th century [Olgeirsson 1957; Gurevich 1972; Steblin-Kamenskij 1984]) which could not yet be denoted with any commonly accepted terms and whose own self-designations are often too complex (like Kazachye Voysko) to have any chance to get transformed into general terms.
So what are our preliminary conclusions at this point? Of course, the socioevolutionary sequences characterized by the simultaneous growth of cultural complexity and non-egalitarianism are rather typical for the medium-range societies. Indeed, with the transition from the low to medium levels of cultural complexity the proportion of egalitarianally organized polities seem to decrease significantly. However, at each level of cultural complexity it appears to be still possible to find more or less egalitarian communities. What is more, even for the medium-range societies we can find a considerable number of documented cases when the growth of cultural complexity went hand in hand with the growth of the egalitarianism (many Greek poleis in the first half of the 1st millennium BC; the Roman civitas in the 6th – 4th centuries BC; North-East Yemen in the second half of the 1st millennium AD – the first half of the 2nd millennium AD; many Swiss and Daghestani communities in the first half of the 2nd millennium AD, etc.); whereas in the 19th – 20th centuries among the most complex societies the proportion of the egalitarianally organized societies increased significantly.
Hence, we think that the egalitarian vs. non-egalitarian human societies should be considered not as stages, but rather as alternatives of social evolution. On the other hand, it justifies the possibility of comparing the correlation matrix of egalitarianism/non-egalitarianism characteristics for the medium and high complexity cultures (see Tables 6 and 7).
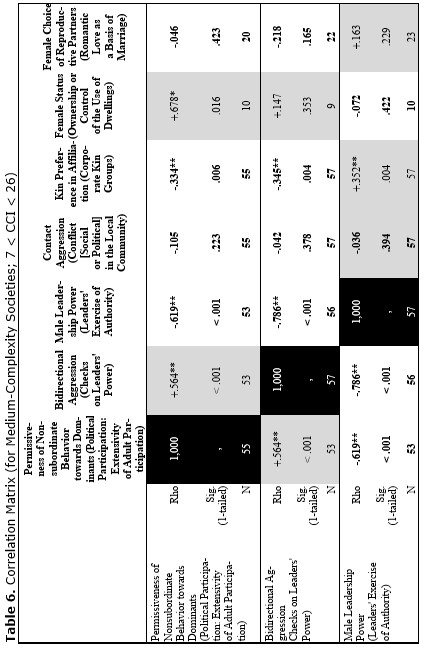
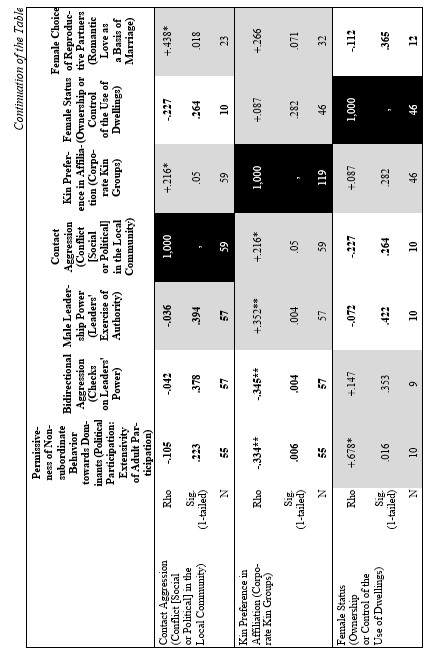
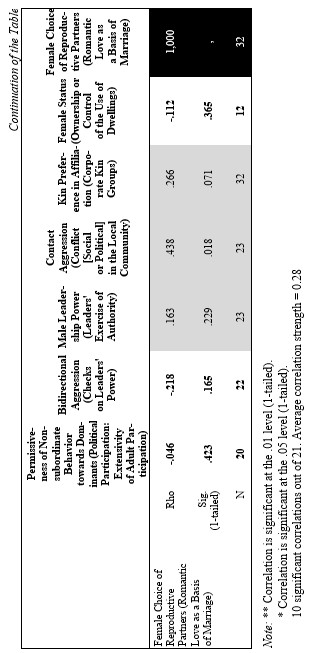
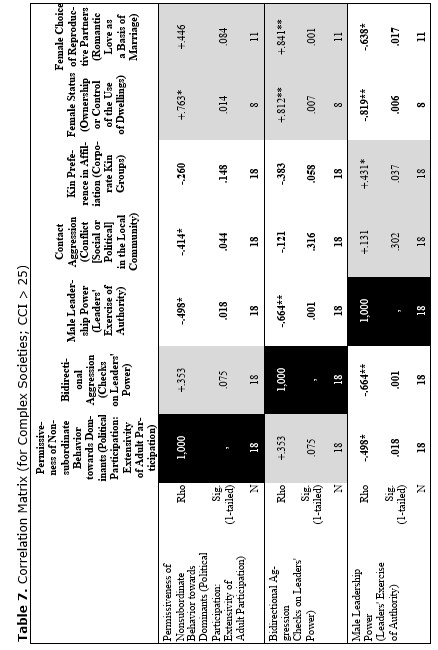
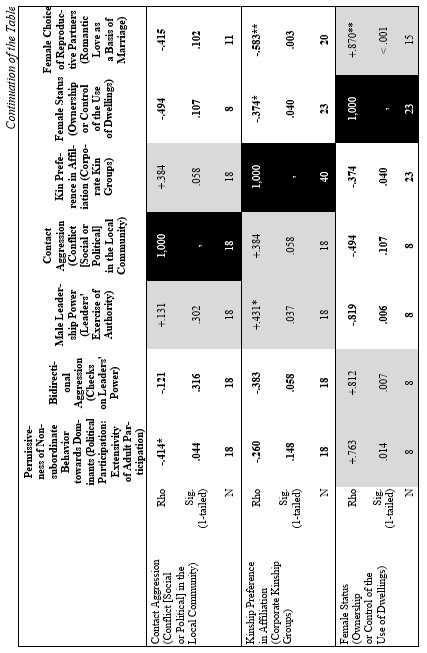
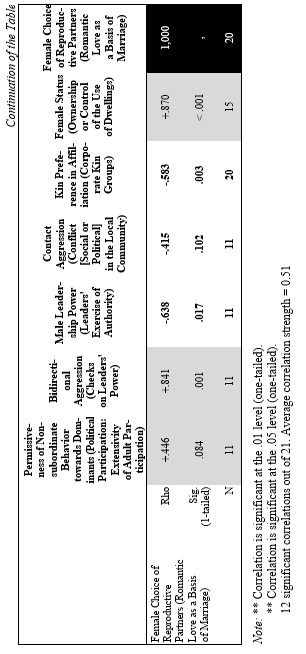
The tables suggest that indeed the transition from the simple cultures to the medium-complexity ones is accompanied by dramatic changes with respect to the relationships between the egalitarianism/non-egalitarianism characteristics. The correlation matrix for the simple hunter-gatherers looked almost identical with the one for the non-human primates. It appears impossible to say the same with respect to the medium-range cultures. The correlation matrix starts to differ. The number of significant correlations drops from 16 to 10, whereas the average correlation strength drops from 0.62 to 0.28. However, what seems even more important is that the whole structure of the correlation matrix changes. In seven cases the sign of the correlation changes to the opposite, whereas in one case a strong and significant negative correlation is replaced with a strong and significant positive correlation.
On the one hand, the disintegration of the ‘primate’ correlation matrix for the medium-range societies might be caused to some extent by the imperfection of measurement of certain variables. It seems most relevant for the measurement of contact aggression which we have used. The standard cross-cultural sample Variable 767 only measures the aggression within communities. However, for the medium-range societies this might not always be the most important type of internal aggression. Indeed, sometimes some non-egalitarian agrarian polities succeeded quite well in the monopolization of the means of aggression, thus imposing relative peace within the communities which they controlled. But this did not mean that the commoners were safe from the contact aggression as a result of this. Sometimes it could mean that the supra-communal authorities would transform themselves into the main contact aggression agents. And this would become quite clear, for example, at time of tax-collection, which could be illustrated by the following citations from original sources:
A scribe moors to a (river-)bank. He reaps the harvest. His retainers are with sticks; the Cushites [retainers of southern origin] are with palm-leaf fibre [whips]. They [say to a peasant]: ‘You, give the barley!’ There is no [barley]. They beat him mercilessly. He is bounded and thrown into a well. He is immersed into water with his hand down, his wife is bounded up in front of him, his children are bound (Papyrus Sallier I, 3 [Berlev 1963: 138]; Egypt, early 1st millennium BC).
I saw those who were led by the guardians to jail and to the Taxation Office [Diwa:n]. One who was brought there was put into the hell-like heat of the midday, so that his head became similar to a boiling pot. His arms were bounded by ropes which cut his joints. He was hung on a wall as if he was a clay pot for cool water in a house. And they beat him at the back of his head as if they beat a drum, one time showing malicious joy with their eyes, and another time pretending to be his friends... And when he begged them to save him from the sun's rays, the tax-collectors kicked him with their feet in response. And meanwhile a jailer poured oil on him, so he was completely covered with a red crust. It lasted until he became exhausted and he could not resist their demands... After that he paid everything that was demanded from him and went away... Then the guardians who torched him came to him and asked more money, as if they were sure that they got him to the state of the complete submissiveness. And if he was slow to answer them, they stripped his turban off his head and injured his neck and his head (Ibn al-Mu`tazz [AD 861–908] 1961: 494; Iraq, late 9th century AD).
Thus, it is quite clear that even the members of the communities ‘pacified’ by non-egalitarian polities could still be subject to the most explicit systematic contact aggression from outside their communities (but still within their societies). Incidentally, this aggression would involve the most severe risk of injuries on the part of commoners. Hence, such non-egalitarian societies would not be so different from the non-egalitarian societies of the non-human primates in this respect, though the absence of the relevant coded data does not permit to detect this.
However, this does not seem to be true with respect to some other egalitarian characteristics, first of all with respect to the low kin preference in affiliation and the high female status. In some cases the overall ‘primate’ correlation matrix appears to work for the medium-range egalitarian cultures for these characteristic too. For example, the egalitarianization of the Classical Greek poleis and the Roman civitas seems to have gone hand in hand with the declining importance of the kinship ties and with concomitant growth of the non-kinship ones (e.g., Berent 1994, 2000; Dozhdev 2000). However, this does not appear to have been accompanied, for example, by a significant growth of the female status. In the highland Daghestan the female status in egalitarian jamâ`âts was a bit higher than in non-egalitarian ones (personal communication with Khizrieva in 2000), but this does not seem possible to maintain with respect to the kinship ties (Aglarov 2014).
However, the complex
societies seem to return to the ‘normal’ primate correlation matrix. Their matrix
structure is once more identical with the one of the non-human primates (and simple
hunter-gatherers). The number of significant correlations and the average correlation
strength also start approaching the ones for the simple hunter-gatherers. It is
no surprise that when analyzing a world-wide sample consisting of the most simple
and most complex cultures (thus omitting the medium-complexity societies) we obtain
a perfect ‘primate’ correlation matrix (see Table 8). All the correlations are in
the predicted direction. Though its average correlation strength is lower than for
both the non-human primates and the simple hunter-gatherers, the number of significant
correlation is even higher than for the non-human primates – the maximum 21 of 21.
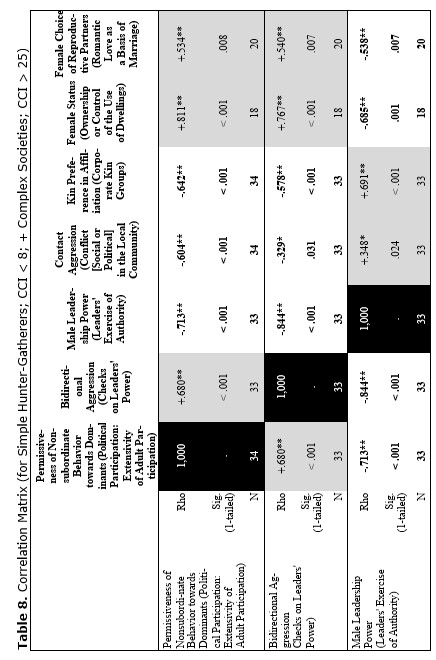
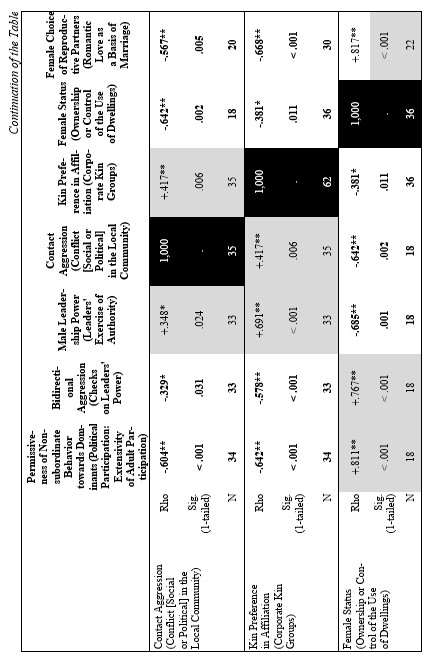
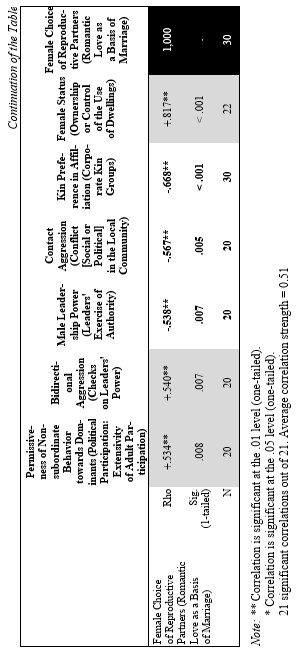
Discussion and Conclusions
To summarize, social interactions in primates are supposed to be strongly structured by dominance and kinship. To understand the reasons of species and even population differences in the types of hierarchies, dispersal patterns and relations within and between sexes both socioecological and of phylogenetic models are needed.
The statistical analysis of our primatological database (see Table 1 above) has shown that the egalitarian type of social organization in primates is associated with the following traits:
1) absence or low incidence of submission demonstration;
2) permissiveness of non-subordinate behavior towards dominants;
3) low level of male leadership power (i.e., the situation when group members systematically dispute the leader);
4) permissiveness on feeding sites;
5) low intensity of contact aggression (though sometimes high frequency);
6) low risk of injury;
7) low risk of severe injury;
8) high level of bidirectional aggression;
9) low level of redirected aggression;
10) low level of female–female intergroup aggression;
11) low level of male–male intergroup aggression;
12) presence of formal biting;
13) support in aggression on the side of victims rather than aggressor;
14) high level of affiliative support;
15) low level of aggressive support;
16) high level of reconciliation after conflict;
17) low kin preferences in reconciliation;
18) low or absent kin preferences in affiliation;
19) high probability of female carrying other females' infants (i.e., permissiveness in access to infants for other females, non-kin included);
20) low or absent kin effect on spatial distribution in feeding;
21) presence of social play between males;
22) high level of male affiliative interaction with juveniles;
23) high level of male–male tolerance (and late emigration of adolescent males in case of female phylopatric species);
24) high level of grooming between males;
25) high level of grooming between non-kin females;
26) the absence of connection between grooming received and dominant status;
27) high female choice of reproductive partner;
28) high female status;
29) different degrees of egalitarianism within males vs. females (in some cases);
30) presence of male attachments with anestrus females;
31) absence of seasonality of reproduction.
The non-egalitarian primate social systems are characterized by generally the same set of parameters with opposite values.
For us one of the most impressive results of our further statistical tests was that the ‘primate’ correlation matrix for the egalitarian vs. non-egalitarian characteristics, while being completely valid for the simple hunter-gatherer cultures, almost disintegrated for the medium-range societies but resurfaced again among the most complex cultures. How could that be explained?
First, let us return to the question of what could account for the near-disintegration of the egalitarianism vs. non-egalitarianism correlation matrix for the medium-range societies.
The immediate explanation seems to be as follows.
It seems possible to find within the medium-range cultures ‘perfectly non-egalitarian’ societies, i.e. societies having almost all the ‘non-egalitarian’ characteristics. However, as was mentioned above, within the medium-range societies we could hardly find any ‘perfectly egalitarian’ cultures, any cultures possessing all the main egalitarian traits, i.e. not only ‘weakly linear and shallow dominance relations’, but also such traits as, for example, low level of contact aggression, high female status (including female choice of reproductive partners), and relatively high importance of non-kinship affiliative ties.
Yet, such societies seem to be found among the most complex cultures. Indeed, most communities in such societies as Britain, the Netherlands, Scandinavian states, etc. seem to possess almost all the characteristics correlating with the egalitarian organization among the primates, i.e. near-absence of Demonstration of Submission, presence of Permissiveness of Non-subordinate Behavior towards Dominants, low levels of Male Leadership Power, Contact Aggression, Risk of Injury, and Severe Injuries, support of a victim rather than an aggressor in the situations of aggression, high levels of Affiliative Support, and low levels of Aggressive Support, high levels of Bidirectional Aggression (possibility of aggression [mainly non-contact, of course, in our case] up the hierarchy), and Reconciliation after Conflict, relatively low levels of Kin Preference in Support in Aggression, Kin Preference in Reconciliation, Kin Preference in Affiliation, high levels of Male–Male Tolerance (for humans for males with different status), Male Affiliative Interaction with Juveniles, Female Carrying Other Females' Infants, relatively low levels of Redirected Aggression, relatively high level of Affiliation between Males (for males with different status), and Social Play between Adult Males, low level of Male–Male Intergroup Aggression, high levels of Male Attachments with Anestrus Females (~ ‘romantic love’), Female Choice of Reproductive Partners and Female Status.
One group of variables (due to obvious reasons) turns out to be irrelevant (Direction of Grooming up the Hierarchy, Grooming between Males, Grooming between Females, Formal Biting, Seasonality of Reproduction). And the only conspicuous exception appears to be Permissiveness on Feeding Sites (supposedly corresponding for the Humans to the presence of privileges in distribution of food). However, as we will see below, even in this respect the difference between the post-industrial and hunter-gatherer egalitarian societies turns out to be not as absolute, as one could imagine.
It appears to be reasonable to view this against the following background. Is this the only group of variables according to which the most complex cultures turn out to be closer to the simplest societies than to the medium-range ones? Of course, not.
The list of such variables is quite long. It includes such important characteristics as the family size (Nimkoff and Middleton 1960; Osmond 1969; Blumberg and Winch 1972; McNett 1973; Pasternak, Ember, and Ember 1997: 235; Ember and Ember 1999: 330; Bondarenko and Korotayev 2000, 2004), polygyny (White and Burton 1988; White 1988; Korotayev and Bondarenko 2000; Bondarenko and Korotayev 2004), male initiations (Precourt 1975; Schlegel and Barry 1980), treatment of aged (Lee and Kezis 1979; Balkwell and Balswick 1981; Lee 1984; Ishii-Kuntz and Lee 1987), unilineal descent organization (Aberle 1961; Service 1971[1962]; Coult and Habenstein 1965; Pasternak 1976; Pasternak, Ember, and Ember 1997: 262; Korotayev 2003a, 2003c, 2004, etc.), corporate kin groups (unilineal and ambilineal) in general (Murdock and Provost 1973), kinship terminology (Korotayev 1999, 2004; Korotayev et al. 2019), communal democracy (Bondarenko and Korotayev 2000; Korotayev 2003a, 2003c), etc.
Many egalitarian hunter-gatherers turn out to be closer to the most complex industrial societies than to the traditional medium-complexity agricultural societies according to such fundamental characteristics as the average life expectancy and the food patterns. Indeed, starting with the Neolithic revolution and till the final formation of the agricultural high civilization an overall steady trend to the decline of the average life expectancy seems to be observed. And this comes as no surprise. Of course, when anthropologists discovered that the Kalahari Bushmen had the food patterns (the kilocalories quantity, proportion and absolute quantities of the proteins, fats, carbohydrates, microelements) conforming well to the World Health Organization recommendations, they seem to have been surprised (Lee and DeVore 1968: 5; Sahlins 1972; Cohen 1977: 39; 1987, 1989, 1995, 1998, 2009; Cohen and Armelagos 1984; Cohen and Crane-Kramer 2007; Ember and Ember 1999: 152–153). However, it is not really surprising. Indeed, the human biogram formed within the conditions of the hunter-gatherer way of life; and it got adapted precisely to those conditions.
The transition to the food production almost inevitably led to the deterioration of the basic conditions of human existence (that is why the foragers do not appear to shift to the food production in absence of foraging economy crisis). Indeed, what is the food production? This is the artificial augmentation of the biomass of a very limited number of species of domesticated plants and animals to the detriment of the biomass of all the other species of edible plants and animals. What is more, the further intensification of agriculture normally led just to the further increase in the contribution to the human diet of more and more limited number of plant species (the most productive and calorigenic ones, and thus hardly containing anything in addition to carbohydrates) to the detriment of all the rest. The diet becomes more and more monotonous, carbohydratic, containing less and less proteins, vitamins, microelements. The growing fixity of settlement leads to the worsening of the sanitary conditions which is further aggravated by the population concentrations. This is accompanied by the growing number of parasites, pathogens which is further aggravated by the development of communications that leads to the spread of more and more diseases in the belt of the high civilizations. As a result we have such figures of average life expectancy (taking into account the extremely high infant mortality) as 18–19 years in Teotihuacan, or 15–16 in Imperial Rome of the Principate Age (Alekseev 1989; Cohen 1977, 1987, 1989, 1995, 1998; Cohen and Armelagos 1984; Fedosova 1994; Kozintsev 1980; Storey 1985).
In some areas of Eurasia this trend seems to have started to get reversed more than 1,000 years ago (and not only in Europe – in the 18th century the highest life expectancy within the high civilization belt seems to have already been attested in Japan [Mel'jantsev 1996: 145]). However, a radical reversal of the above-mentioned trend was only achieved in the 19th and 20th centuries (see e.g., Caldwell 2006; Livi-Bacci 2012). It seems necessary to emphasize that it was achieved to a considerable extent through a virtual unconscious artificial imitation of many fundamental characteristics and life conditions of the Paleolithic hunter-gatherers. The diet of the populations of the modern egalitarian societies (which are characterized by the highest average life expectancy) in an unconscious way artificially imitates to a considerable extent that of an Upper Paleolithic hunter-gatherer[6] – this diet is characterized by heightened quantities of proteins, vitamins, reduced quantities of carbohydrates, fats, etc. We also observe the unconscious artificial imitation of many other life conditions of a hunter-gatherer; for example the naturally better (in comparison with an average pre-industrial agriculturist) hygienic situation is artificially ‘imitated’; even the typical physical load of a hunter is artificially imitated in an unconscious way through the physical exercises. The question arises whether it is possible to add to this list a set of egalitarian behaviors (typical of egalitarian hunter-gatherers).
However, the same problem could be also viewed in a rather different (but highly complementary) perspective. It seems important to pay special attention to the socialization patterns. Some of their characteristics among the egalitarian hunter-gatherers are again closer to the ones in the developed industrial and post-industrial egalitarian cultures rather than among the non-egalitarian traditional agriculturists. Indeed, the traditional non-egalitarian agriculturists put emphasis on the dependence training, whereas both the egalitarian hunter-gatherers and the developed industrial and post-industrial egalitarian cultures practice the independence training (Barry et al. 1959; Dawson 1967; Berry 1971, 1976). And there is nothing strange about it notwithstanding the point that it seems difficult to imagine anything more different than the post-industrial societies compared with the simple hunter-gatherers. Indeed, in spite of all the immense differences between them, they appear to have basically similar fundamental conditions and characteristics of existence, which are, on the other hand, significantly different from the ones which are typical for traditional agricultural societies. The traditional agriculturists live in accordance with more or less predictable natural cycles which make the following of the traditional routines quite adaptive. On the other hand, the fundamental feature, which is typical for both the egalitarian hunter-gatherers and the egalitarian post-industrial societies, is the basic unpredictability of the constantly changing (and not controlled by individual persons) fundamental conditions of production and reproduction. This makes the following of the traditional routines and blind submission to the traditional authorities counter-productive, this makes it necessary to constantly take independent decisions.
Let us take, for example, the !Kung Bushmen conditions of production and reproduction. The Nyae Nyae region, where the !Kung Bushmen studied by Marshall and Lee lived, is characterized by highly imbalanced (both in terms of time and space) regime of precipitation (Leistner 1979: 110–122; Cole 1966: 55; Gould 1991). The bulk of rains come in January – March mostly in the form of monsoon torrential storms. Even in usual years some parts of the region may be literally flooded whereas the others receive only sprinkle of rain (Marshall 1973: 108; 1976: 2–7, 62–73). This situation strongly affects the system of residence among the Nyae Nyae Bushmen. In a desperate situation a hunter and his family may have to leave his basic hunting grounds (n!ore) and seek refuge in the area where the rains were normal or where there is a plentiful permanent source of water (well) and where his relatives either on paternal or maternal side or relatives of his wife live. The residence of the Nyae Nyae !Kung is thus, of necessity opportunistic (Idem. 1973: 108; 1976: 94). Quite predictably no unilineal descent groups with unilocal residence are observed among them (Idem. 1976: 380–411).[7]
According to B. Pasternak, C. Ember and M. Ember,
Descent groups lose viability in complex state-organized, commercial-industrial societies... In complex societies, it is individuals (not families or larger kin groups) who take advantage of economic or occupational opportunities; when someone moves to a new job parents and siblings are not likely to go along (and cousins and aunts and uncles even less likely) (Pasternak, Ember, and Ember 1997: 263).
Thus, we observe how substantially similar (though formally different) conditions produce similar outcomes. For the Bushmen it is not predictable where the rainfall will be available next year. Within such a situation, for example, the unilocal marital residence would be explicitly maladaptive. It would be rather unproductive for married couples to settle constantly with, say, the husbands' paternal relatives because a better situation might be available in quite a different place. Within the above described situation the residence should be necessarily opportunistic in order to be adaptive which would inevitably inhibit the formation of the unilineal descent groups. But just the same (substantially) is observed in the post-industrial egalitarian societies. One cannot be sure that particular jobs will be still available in the given locality in ten or even five years, or in a year. Within the traditional agrarian society for, say, a son of a village smith an option to learn the profession of his father, to bring his wife to his father's place and to live just doing copiously the same which his father did would be rather adaptive and risk-reducing. It is no more in commercial-industrial (let alone post-industrial) societies. Like a Bushman (or a Pitjantjara) ‘hunting a rain’, a member of egalitarian post-industrial society has to ‘hunter a job’. The fact that the rainfall was available in a given Kalahari (or Australian West Desert) locality this year, does not mean that it will be next year. The fact that there was demand in your profession in this locality this year does not mean that this demand will remain in ten years, in five or even next year.[8] In both cases the situation constantly demands from the individuals to make independent decisions. Both situations make flexible egalitarian structures highly adaptive. Thus, the essentially similar (though formally different) conditions of production and reproduction stimulate in the Homo sapiens populations the development of similar basic social structures (and similar sets of behavioral patterns).
Homo sapiens is not the only primate species whose groups living in different conditions can develop different social structures and behavioral patterns which suggests that at least some primate species could have more than one set of behavioral predispositions whereas different sets would be activated in different conditions (e.g., Williamson and Dunbar 1999; Butovskaya and Kozintsev 1996b). Homo sapiens seems to fall directly into this category. It appears that the Homo sapiens primates have both egalitarian and non-egalitarian sets of behavioral predisposition. For the hunter-gatherer the complete egalitarian set of behavioral patterns seems to have developed mainly in extreme ecological conditions, where this set appears to have increased significantly the survival chances of the respective populations. Hence, the presence of two sets of behavioral predispositions in the human biogram appears to have been quite adaptive within the conditions of hunter-gatherer mode of life within which that biogram was formed.
Paradoxically, the set of behavioral predispositions, which was highly adaptive for the simple hunter-gatherers in harsh and unpredictable environment seems to have become highly adaptive again within the formally different but substantively similar conditions of the developed commercial-industrial and post-industrial societies.
The results of the present study confirm our initial suggestion that the behavioral patterns of non-human and human primates are quite comparable and some general paradigms are applicable in both cases. The study of non-human primate social organization can be useful in the development of an effective working model for understanding the process of differentiation of dominance styles in different human cultures.
Acknowledgements
This paper was prepared in accordance with the state assignment of the Institute of Ethnology and Anthropology RAS (MB). We would also like to express our gratitude to Professor Nikolay Kradin (Dept. of Anthropology, Far East Technical University, Vladivostok) for useful references regarding complex egalitarian polities of the Cossacks and Early Medieval Icelanders.
References
Abegg C., Thierry B., and Kaumanns W. 1996. Reconciliation in Two Groups of Lion-Tailed Macaques (Macaca silenus). Int. J. Primatol. 17: 803–816.
Aberle D. F. 1961. Matrilineal Descent in Cross-Cultural Perspective. Matrilineal Kinship / Ed. by D. M. Schneider, and K. Gough, pp. 655–727. Berkeley: University of California Press.
Abu Ghanim F. A. A. 1985. Al-bunyah al-qabaliyyah fî 'l-Yaman bayna 'l-istimrâr wa-'l-taghayyur [The Tribal Structure in Yemen between Continuity and Change]. Dimashq: al-Kâtib al-`Arabiyy.
Abu Ghanim F. A. A. 1990. Al-qabîlah wa-'l-dawlah fi 'l-Yaman [The Tribe and the State in Yemen]. al-Qâhira: Dâr al-Manâr.
Adams R. N. 1975. Energy and Structure. Austin – London: University of Texas Press.
Aglarov M. A. 2014. Village Community in Highland Daghestan in the 17th to early 19th Centuries. Makhachkala: Nauka – Daghestan. In Russian (Агларов М. А. Сельская община в Нагорном Дагестане в XVII – начале XIX в. Махачкала: Наука – Дагестан).
Azumagakito T., Suzuki R., and Arita T. 2018. An Integrated Model of Gene-Culture Coevolution of Language Mediated by Phenotypic Plasticity. Scientific Reports 8(1): 1–11.
Alekseev V. P. 1989. Paleodemography:
Contents and Results. Historical Demography: Problems, Judgments, Tasks / Ed. by Yu. A. Poljakov, pp. 20–32. Moscow: Nauka. In Russian (Алексеев В. П.
Палеодемография: cодержание и результаты. Историческая демография: проблемы,
суждения, задачи / Ред. Ю. А. Поляков,
с. 20–32. М.: Наука).
Amici F., Call J., Watzek J., Brosnan S., and Aureli F. 2018. Social Inhibition and Behavioural Flexibility When the Context Changes: A Comparison Across Six Primate Species. Scientific Reports 8(1): 1–9.
Ammar H. 1954. Growing up in an Egyptian Village. London: Routledge & Kegan Paul.
Artemova O. Yu. 1987. Personality and Social Norms in the Archaic Community. Moscow: Nauka. In Russian (Артемова О. Ю. Личность и социальные нормы в раннепервобытной общине. M.: Наука).
Artemova O. Yu. 1989. On the Problem of the Primitive Egalitarianism. Linguistic Reconstruction and the Most Ancient History of the East / Ed. by S. Blümchen et al. Part 3, pp. 3–5. Moscow: Nauka. In Russian (Артемова О. Ю. О проблеме примитивного эгалитаризма. Лингвистическая реконструкция и древнейшая история Востока / Ред. С. И. Блюмхен и др., Ч. 3., c. 3–5. М.: Наука).
Artemova O. Yu. 1991. Egalitarian and Non-egalitarian Primitive Societies. Archaic Society: Basic Problems of Evolutionary Sociology. Vol. 1 / Ed. by A. Korotayev and V. Chubarov, pp. 44–91. Moscow: Institute of Russian History, Soviet Academy of Sciences. In Russian (Артемова О. Ю. Эгалитарные и неэгалитарные первобытные общества. Архаическое общество: узловые проблемы социологии развития / Ред. А. В. Коротаев и В. В. Чубаров. Т. I., c. 44–91. М.: Институт истории СССР АН СССР).
Artemova O. Yu. 1993. Primitive Egalitarianism and Early Forms of Social Differentiation. Early Forms of Social Stratification: Genesis, Historical Dynamics, Testamentary-Political Functions. Dedicated to L. E. Kubbel / Ed. by V. A. Popov, pp. 40–70. Moscow: Vostochnaya Literatura. In Russian (Артемова О. Ю. Первобытный эгалитаризм и ранние формы социальной дифференциации. Ранние формы социальной стратификации: генезис, историческая динамика, потестарно-политические функции. Памяти Л. Е. Куббеля / Ред. В. А. Попов, c. 40–70. М.: Восточная литература).
Artemova O. Yu. 2000. Monopolization of Information and Social Inequality. Alternatives of Social Evolution / Ed. by N. N. Kradin, A. V. Korotayev, D. M. Bondarenko, V. de Munck, and P. Wason, pp. 1–12. Vladivostok: Dal'nauka.
Artemova O. Yu. 2003. Monopolization of Knowledge, Social Inequality, and Female Status: A Cross-Cultural Study. Cross-Cultural Research 37(1): 62–80.
Artemova O. Yu. 2009. The Kin of Esau: Hunters, Gatherers, Fishers. Moscow: Smysl. In Russian (Артемова О. Ю. Колено Исава. Охотники, собиратели, рыболовы. М.: Смысл).
Artemova O. Yu. 2016. Monopolization of Knowledge, Social Inequality and Egalitarianism. Hunter Gatherer Research 2: 1–36.
Artemova O. Yu. 2020. Equality as a Human Categorical Imperative. Prehistoric Archaeology. Journal of Interdisciplinary Studies 1: 64–91.
Aureli F. 1992. Post-Conflict Behavior Among Wild Long-Tailed Macaques (Macaca fascicularis). Behav. Ecol. Sociobiol. 31: 329–337.
Aureli F., and van Schaik C. P. 1991. Post-Conflict Behavior in Long-Tailed Macaques (Macaca fascicularis). I. The Social Events. Ethology 89: 89–100.
Aureli F., van Schaik C. P., and van Hooff J. A. 1989. Functional Aspects of Reconciliation Among Captive Long-Tailed Macaques (Macaca fascicularis). Amer. J. of Primatol. 19: 39–51.
Aureli F., Das M., Verleur D., and van Hooff J. A. 1994. Post-conflict Social Interactions Among Barbary Macaques (Macaca sylvanus). Int. J. Primatol. 15(3): 471–485.
Aureli F., Veenema C., van Panthaleon C. J., van Eck, and van Hooff J. A. 1993. Reconciliation, Consolation, and Redirection in Japanese Macaques (Macaca fuscata). Behavior 124: 1–21.
Bailey G., and Peoples J. 1999. Introduction to Cultural Anthropology. Belmont, CA: Wadsworth.
Baines T. 1864. Explorations in South-West Africa. London: Longman.
Balasubramaniam K. N., Dittmar K., Berman C. M., Butovskaya M., Cooper M. A., Majolo B. et al. 2012. Hierarchical Steepness and Phylogenetic Models: Phylogenetic Signals in Macaca. Animal Behaviour 83(5): 1207–1218.
Balasubramaniam K. N., Beisner B. A., Berman C. M., De Marco A., Duboscq J., Koirala S. et al. 2018. The Influence of Phylogeny, Social Style, and Sociodemographic Factors on Macaque Social Network Structure. American Journal of Primatology 80(1): e22727.
Balkwell C., and Balswick J. 1981. Subsistence Economy, Family Structure and the Status of the Elderly. Journal of Marriage and Family 43: 423–429.
Barnard A. 1992. Hunters and Herders of Southern Africa: A Comparative Ethnography of the Khoisan Peoples. Cambridge: Cambridge University Press.
Barry H. III, Child I. L., and Bacon M. K. 1959. Relation of Child Training to Subsistence Economy. American Anthropologist 61: 51–63.
Barton R. F. 1919. Ifugao Law. Berkeley, CA: University of California Press (University of California Publications in American Archaeology and Ethnology, XV).
Barton R. F. 1922. Ifugao Economics. Berkeley, CA: University of California Press (University of California Publications in American Archaeology and Ethnology, XV/5).
Barton R. F. 1938. Philippine Pagans. London: Routledge.
Barton R. A. 1993. Sociospatial Mechanisms of Feeding Competition in Female Olive Baboons, Papio Anubis. Anim. Behav. 46: 791–802.
Barton R. A., Byrne R. W., and Whiten A. 1996. Ecology, Feeding Competition and Social Structure in Baboons. Behav. Ecol. Sociobiol. 38: 321–329.
Basedow H. 1913. Notes on the Natives of Bathurst Island, North Australia. Journal of the Royal Anthropological Institute 43: 291–323.
Berent M. 1994. Stateless Polis. Unpublished PhD. thesis manuscript. Cambridge, UK.
Berent M. 1996. Hobbes and the ‘Greek Tongues’. History of Political Thought 17: 36–59.
Berent M. 1998. Stasis, or the Greek Invention of Politics. History of Political Thought 19: 331–362.
Berent M. 2000. The Stateless Polis. Alternatives
of Social Evolution / Ed. by
N. N. Kradin, A. V. Korotayev, V. de Munck, V. A. Lynsha, D. M. Bondarenko, and
P. Wason, pp. 225–241. Vladivostok: Dal'nauka.
Berent M. 2004. Greece: The Stateless Polis (the 11th – the 4th Centuries B. C.). The Early State, Its Alternatives and Analogues / Ed. by L. E. Grinin, R. L. Carneiro, D. M. Bondarenko, N. N. Kradin, and A. V. Korotayev, pp. 364–387. Volgograd: Uchitel.
Berent M. 2006. The Stateless Polis: A Reply to Critics. Social Evolution & History 5(1): 141–163.
Berezkin Yu. E. 1994. The Apa Tanis and the Ancient Near East. Alternative Model of Complex Society. Kunstkamera: Etnograficheskiye Tetradi 4: 5–19. In Russian (Березкин Ю. Е. Апатани и древнейший Восток: альтернативная модель сложного общества. Кунсткамера: этнографические тетради 4: 5–19).
Berezkin Yu. E. 1995a. Chiefdoms and Acephalic Complex Societies: Archaeological Data and Ethnographic Parallels. Early Forms of Political Organization: From Primitivity to Statehood / Ed. by V. A. Popov, pp. 62–78. Moscow: Vostochnaya literatura. In Russian (Березкин Ю. Е. Вождества и акефальные сложные общества: данные археологии и этнографические параллели. Ранние формы политической организации / Ред. В. А. Попов, с. 62–78. М.: Восточная литература).
Berezkin Yu. E. 1995b. Alternative Models of Middle Range Society. ‘Individualistic’
Asia vs. ‘Collectivistic’ America? Alternative Pathways to Early State /
Ed. by
N. N. Kradin, and V. A. Lynsha, pp. 75–83. Vladivostok: Dal'nauka.
Berezkin Yu. E. 1997. America and the Middle East. The Forms of Sociopolitical Organization in the Pre-State Period. Vestnik drevnei istorii 2: 3–24. In Russian (Березкин Ю. Е. Америка и Ближний Восток: формы социополитической организации в догосударственную эпоху. Вестник древней истории 2: 3–24).
Berezkin Yu. E. 2000. Once Again on Horizontal and Vertical
Links in Structure of the Middle Range Societies. Alternatives of Social Evolution
/ Ed. by N. N. Kradin,
A. V. Korotayev, V. de Munck, V. A. Lynsha, D. M. Bondarenko, and P. Wason, pp.
220–224. Vladivostok: Dal'nauka.
Berlev O. D. 1963. Papyrus Sallier I. Chrestomathy on the History of Ancient East / Ed. by V. V. Struve, and D. G. Reder, pp. 137–138. Mosсow: Isdatel'stvo vostochnoj literatury. In Russian (Берлев О. Д. Папирус Sallier I. Хрестоматия по истории Древнего Востока / Ред. В. В. Струве, Д. Г. Редер, с. 137–138. М.: Издательство восточной литературы).
Berndt R. 1942–1943. A Preliminary Report of Field Work in the Ooldea Region, Western South Australia. Oceania 13(1): 51–70; (2): 143–169; (3): 243–280; (4): 239–275.
Berndt R. M., and Berndt C. H. 1964. The World of the First Australians. Chicago: University of Chicago Press.
Berndt R. M., and Berndt C. H. 1970. Man, Land, and Myth in North Australia: The Gunwinggu People. East Lansing, MI: Michigan State University.
Berry J. W. 1971. Ecological and Cultural Factors in Spatial Perceptual Development. Canadian Journal of Behavioral Science 3: 324–336.
Berry J. W. 1976. Human Ecology and Cognitive Style. New York: Wiley.
Bertrand M. 1969. The Behavioral Repertoire of the Stumptail Macaque. Basel: S. Barger.
Betzig L. 1997. Human Nature: A Critical Reader. New York, NY: Oxford University Press.
Bjerre J. 1960. The Kalahari. New York: Hill & Wang.
Bjerre J. 1963. The Lost World of Kalahari. Moscow: Geografgiz. In Russian (Бьерре Й. Затерянный мир Калахари. М.: Географгиз).
Blainey G. 1975. Triumph of the Nomads: A History of Ancient Australia. Melbourne – Sydney: MacMillan Company of Australia.
Bleek D. F. 1928. The Naron. Cambridge: Cambridge University Press.
Bleek D. F. 1931. The Hadzapi or Watindiga of Tanganyika Territory. Africa 4: 273–286.
Blumberg R. L., and Winch R. F. 1972. Societal Complexity and Familial Complexity: Evidence for the Curvilinear Hypothesis. American Journal of Sociology 77: 898–920.
Blurton J. N., Hawkes K., and O'Connell J. F. 1999. Some Current Ideas about the Evolution of the Human Life History. Comparative Primate Socioecology / Ed. by P. C. Lee, pp. 140–167. Cambridge: Cambridge Univ. Press.
Bondarenko D. M. 2007. Homoarchy as a Principle of Sociopolitical Organization. Anthropos 102: 187–199.
Bondarenko D. M. 2008. Kinship, Territoriality and the Early State Lower Limit. Social Evolution and History 7(1): 19–53.
Bondarenko D. M. 2014. On the Nature and Features of the (Early) State: An Anthropological Reanalysis. Zeitschrift für Ethnologie 139(2): 215–232.
Bondarenko D. M. 2020. Social Institutions and Basic Principles of Societal Organization. The Evolution of Social Institutions. Interdisciplinary Perspectives / Ed. by D. M. Bondarenko, S. A. Kowalewski, and D. B. Small, pp. 51–78. Cham: Springer. DOI: 10.1007/978-3-030-51437-2_3.
Bondarenko D. M., Grinin L. E., and Korotayev A. V. 2002. Alternative Pathways of Social Evolution. Social Evolution & History 1(1): 54–79.
Bondarenko D. M., Grinin L. E., and Korotayev A. V. 2004. Alternatives of Social Evolution. The Early State, Its Alternatives and Analogues / Ed. by L. E. Grinin, R. L. Carneiro, D. M. Bondarenko, N. N. Kradin, and A. V. Korotayev, pp. 3–27. Volgograd: Uchitel.
Bondarenko D. M., Grinin L. E., and Korotayev A. V. 2011. Social Evolution: Alternatives and Variations (Introduction). Evolution: Cosmic, Biological, and Social / Ed. by L. E. Grinin, R. L. Carneiro, A. V. Korotayev, and F. Spier, pp. 212–250. Volgograd: ‘Uchitel’ Publishing House.
Bondarenko D. M., and Korotayev A. V. 2000. Family Size and Community Organization: A Cross-Cultural Comparison. Cross-Cultural Research. The Journal of Comparative Social Science 34(2):152–189.
Bondarenko D. M., and Korotayev A. V. 2004. A Historical-Anthropological Look at Some Sociopolitical Problems of Second and Third World Countries. Community, Identity and the State. Comparing Africa, Eurasia, Latin America and the Middle East / Ed. by M. Gammer, pp. 14–41. London – New York: Routledge.
Brandt M. 2017. Tribes and Politics in Yemen: A History of the Houthi Conflict. New York: Oxford University Press.
vom Bruck G. 1993. Réconciliation ambiguë: une perspective anthropologique sur le concept de la violence légitime dans l'imamat du Yémen. La violence et l'état: Formes et évolution d'un monopole / Ed. by E. LeRoy, and T. von Trotha, pp. 85–103. Paris: CNRS.
Buckley W. 1852. The Life and Adventures of William Buckley. Tasmania: Archibald Macdougall.
Buckley W. 1966. Australian Robinzon. Moscow: Nauka. In Russian (Бакли У. Австралийский Робинзон. M.: Наука).
Busse C., and Hamilton W. J. III. 1981. Infant Carrying by Male Chacma Baboons. Science 212: 1281–1283.
Butovskaya M. L. 1993. Kinship and Different Dominance Styles in Three Species of the Genus Macaca (M. arctoides, M. mulatta, and M. fascicularis). Folia Primatologica 60: 210–224.
Butovskaya
M. L. 1995. Some Aspects of Hominid Socioecology According to
Primatological Data. Man and Environment in the Paleolithic / Ed. by H. Ullrich,
pp. 309–316. Liege.
Butovskaya M. L. 1999a. Coping with Social Tension in Primate Societies: Strategic Modelling of Early Hominid Lifestyles. Hominid Evolution: Lifestyles and Survival Strategies / Ed. by H. Ulrich, pp. 44–54. Schwelm: Edition Archaea.
Butovskaya
M. L. 1999b. The Evolution of Human Behavior: The
Relationship Between the Biological and the Social. World Anthropology at the
Turn of the Centuries. Abstracts of the 4th International Congress
of Aleš Hrdlička / Ed. by
D. Sabik, J. Vigner, and M. Vigner, pp. 26–27. Prague: Charles University.
Butovskaya M. L. 2020. Primates as Living Links to Our Past: Variations in Hierarchy Steepness but not Real Egalitarianism. Pervobytnaya Arkheologiya 1: 13–26.
Butovskaya M. L., and Fainberg L. A. 1993. On the Origin of Human Society. Moscow: Nauka.
Butovskaya M. L., and Kozintsev A. G. 1996a. Gender-related Factors Affecting Primate Social Behavior: Grooming, Rank, Age, and Kinship in Bisexual and All-Male Groups of Stumptail Macaques. Asm. J. Phys. Anthropol. 11: 39–54.
Butovskaya M. L., and Kozintsev A. G. 1996b. Group History and Social Style: The Case of Crab-Eating Monkeys. Anthropologie 34: 165–176.
Butovskaya M. L., Kozintsev A. G., Meishvili N. V., and Chalian V. G. 1998. Reconciliation in Hamadryas Baboons. Abstracts of the 17th Intern. Congr. of the Primatological Society, Antananarivo, August 10–14.
Butovskaya M. L., and Rostovtseva V. V. 2021. Evolution of Altruism and Cooperation of Humans: Biosocial Perspective. Moscow: LENAND (In Press). In Russian (Бутовская М. Л., Ростовцева В. В. Эволюция альтруизма и кооперации человека: биосоциальная перспектива. М.: ЛЕНАНД (в печати)).
Caldecott J. O. 1986a. An Ecological and Behavioral Study of the Pig-Tailed Macaques. Basel: Karger.
Caldecott J. O. 1986b. Mating Patterns, Societies and the Ecogeography of Macaques. Anim. Behav. 34: 208–220.
Caldwell J. C. 2006. Demographic Transition Theory. Dordrecht: Springer.
Carneiro R. L. 1970. A Theory of the Origin of the State. Science 169: 733–738.
Carneiro R. L. 1981. The Chiefdom: Precursor of the State. The Transition to Statehood in the New World / Ed. by G. D. Jones, and R. R. Kautz, pp. 37–79. Cambridge: Cambridge University Press.
Carneiro R. L. 1987. Cross-Currents in the Theory of State Formation. American Ethnologist 14: 756–770.
Carneiro R. L. 1991. The Nature of the Chiefdom as Revealed by Evidence from the Cauca Valley of Colombia. Profiles in Cultural Evolution / Ed. by A. T. Rambo, and K. Gillogly, pp. 167–190. Ann Arbour: Museum of Anthropology, University of Michigan (Anthropology Papers, Museum of Anthropology, University of Michigan, No. 85).
Carneiro R. L. 2000. Process vs. Stages: A False Dichotomy in Tracing the Rise of the State. Alternatives of Social Evolution / Ed. by N. N. Kradin, A. V. Korotayev, V. de Munck, V. A. Lynsha, D. M. Bondarenko, and P. Wason, pp. 52–58. Vladivostok: Dal'nauka.
Castles D. L., and Whiten A. 1998a. Post-Conflict
Behavior of Wild Olive Baboons.
I. Reconciliation, Redirection and Consolation. Ethology 104: 126–147.
Castles D. L., and Whiten A. 1998b. Post-Conflict
Behavior of Wild Olive Baboons.
II. Stress and Self-directed Behavior. Ethology
104: 148–160.
Chalian V. G., Meishvili N. V., and Butovskaya M. L. 1997. The Role of Females in the Community of Hamadryas Baboons (Papio Hamadryas). I. Female Dominance Hierarchy in the Gumista Reserve. Vestnik antropologii 3: 126–145. In Russian (Чалян В. Г., Мейшвили Н. В., Бутовская М. Л Роль самок в сообществе павианов гамадрилов (Papio Hamadryas). 1. Иерархия доминирования самок в стаде Гумистинского заказника. Вестник антропологии 3: 126–145).
Chaseling W. 1957. Yulengor. Nomads of Arnhen Land. London: Epworth.
Chelhod J. 1970. L'Organisation sociale au Yémen. L'Ethnographie 64: 61–86.
Chelhod J. 1979. Social Organization in Yemen. Dira:sa:t Yamaniyyah 3: 47–62.
Chelhod J. 1985. Les structures sociales
et familiales. L'Arabie du Sud: histoire et
civilization. 3. Culture et institutions du Yémen / Ed. by J. Chelhod,
pp. 15–123. Paris: Maisonneuve et Larose (Islam d'hier et d'aujourd'hui, 25).
Cheney D. L., and Seyfarth R. M. 1986. The Recognition of Social Alliances by Vervet Monkeys, Cercopithecus aethiops. Anim. Behav. 34: 1722–1731.
Cheney D. L., and Seyfarth R. M. 1989. Redirected Aggression and Reconciliaton Among Vervet Monkeys, Cercopithecus aethiops. Behavior 110: 258–275.
Chirkin V. E. 1955. On the Study of States Which Appeared in the Course of Slave and Peasant Rebellions. Voprosy istorii 9: 23–34. In Russian (Чиркин В. Е. Об изучении государственных образований, возникавших в ходе восстаний рабов и крестьян. Вопросы истории 9: 23–34).
Chism J. B., and Rowell T. E. 1986. Mating and Residence Patterns in Male Patas Monkeys. Ethology 72: 31–39.
Clutton-Brock T. H., and Harvey P. H. 1977. Species Differences in Feeding and Ranging Behavior in Primates. Primate Ecology: Studies of Feeding Behavior in Lemurs, Monkeys and Apes / Ed. by T. H. Clutton-Brock, pp. 557–579. London: Academic Press.
Cohen M. N. 1977. The Food Crisis in Prehistory: Overpopulation and the Origins of Agriculture. New Haven, CT: Yale University Press.
Cohen M. N. 1987. The Significance of Long-Term Changes in Human Diet and Food Economy. Food and Evolution: Toward a Theory of Human Food Habits / Ed. by M. Harris, and E. Ross, pp. 261–283. Philadelphia: Temple University Press.
Cohen M. N. 1989. Health and the Rise of Civilization. New Haven, CT: Yale University Press.
Cohen M. N. 1995. The Osteological Paradox Reconsidered. Current Anthropology 35: 629–631.
Cohen M. N. 1998. Were Early Agriculturalists Less Healthy Than Food Collectors? Research Frontiers in Anthropology / Ed. by C. R. Ember, M. Ember, and P. N. Peregrine, pp. 61–83. Upper Saddle River, NJ: Prentice Hall.
Cohen M. N. 2009. Introduction: Rethinking the Origins of Agriculture. Current Anthropology 50(5): 591–595.
Cohen M. N., and Armelagos G. J. (Eds.) 1984. Paleopathology at the Origins of Agriculture. Orlando, FL: Academic Press.
Cohen M. N., and Crane‐Kramer G. (Eds.) 2007. Bioarchaeological Interpretations of the Human Past: Local, Regional, and Global Perspectives Series. Gainesville, FL: University Press of Florida.
Cole M. M. 1966. South Africa. London: Methuen – New York: Duttan.
Colmenares F., Lozano M. G., and Torres P. 1994. Harem Social
Structure in a Multi-Harem Colony of Baboons (Papio spp.): A Test of the
Hypothesis of the ‘Star-Shaped’ Sociogram. Current Primatology. II. Social
Development, Learning and Behavior / Ed. by J. J. Roeder, B. Thierry, J. R.
Anderson, and N. Herrenshmidt,
pp. 93–101. Strasbourg: University Louse Pasteur.
Colmenares F., and Lozaro-Perea C. 1994. Greeting and Grooming During Social Conflicts in Baboons: Strategic Uses and Social Functions. Current Primatology. II. Social Development, Learning and Behavior / Ed. by J. J. Roeder, B. Thierry, J. R. Anderson, and N. Herrenshmidt, pp. 165–174. Strasbourg: University Louse Pasteur.
Coult A. D., and Habenstein R. W. 1965. Cross Tabulations of Murdock's World Ethnographic Sample. Columbia, MO: University of Missouri Press.
Crumley C. L. 2007. Contextual Constraints on State Structure. Alternativeness in Cultural History: Heterarchy and Homoarchy as Evolutionary Trajectories / Ed. by D. M. Bondarenko, and A. A. Nemirovskiy, pp. 3–22. Moscow: Center for Civilizational and Regional Studies Press.
Daly M., and Wilson M. 1988a. Evolutionary Social Psychology and Family Homicide. Science 242: 519–524.
Daly M., and Wilson M. 1988b. Homicide. New York, NY: Aldine de Gruyter.
Datta S. B. 1989. Demographic Influences on Dominance Structure Among Female Primates. Comparative Socioecology. The Behavioral Ecology of Humans and Other Mammals / Ed. by V. Standen, and R. A. Foley, pp. 265–284. Oxford: Oxford Univ. Press.
Dawson J. L. M. 1967. Cultural and Physiological Influences upon Spatial-Perceptual Processes in West Africa. International Journal of Psychology 2: 115–28, 171–85.
Deag J. M. 1974. A Study of the Social Behavior and Ecology of the Wild Barbary Macaque, Macaca sylvanus L. PhD thesis. Bristol: University of Bristol.
Deng Z., and Zhao Q. 1987. Social Structure in a Wild Group of Macaca thibetana at Mount Emai, China. Folia primatol. 49: 1–10.
Diakonoff I. M. 1983. History of the Ancient East. I. Mesopotamia. Moscow: Nauka. In Russian (Дьяконов И. М. История Древнего Востока. I. Месопотамия. М.: Наука).
Divale W., Khaltourina D., and Korotayev A. 2002. A Corrected Version of the Standard Cross-Cultural Sample Database. World Cultures 13(1): 62–98.
Dozhdev D. V. 2000. Ancient Rome. Civilizational Models of Politogenesis / Ed. by D. M. Bondarenko, and A. V. Korotayev. Moscow: Institut Afriki.
Dostal W. 1974. Sozio-ökonomische Aspekte der Stammesdemokratie in Nordost-Yemen. Sociologus 24: 1–15.
Dostal W. 1985. Egalität und Klassengesellschaft in Südarabien. Anthropologische Untersuchungen zur sozialen Evolution. Horn – Wien: Ferdinand Berger & Söhne (Wiener Beiträge zur Kulturgeschichte und Linguistik, 20).
Dostal W. 1990. Eduard Glaser – Forschungen im Yemen. Wien: Verlag der Österreichischen Akademie der Wissenschaften.
Dozhdev D. V. 2004. Rome: Socio-Political Evolution in the 8th –
2nd Centuries BC. The Early State, Its Alternatives and Analogues
/ Ed. by L. E. Grinin, R. L. Carneiro,
D. M. Bondarenko, N. N. Kradin, and A. V. Korotayev, pp. 388–418. Volgograd:
Uchitel.
Dresch P. 1984a. The Position of Shaykhs among the Northern Tribes of Yemen. Man 19: 31–49.
Dresch P. 1984b. Tribal Relations and Political History in Upper Yemen. Contemporary Yemen: Politics and Historical Background / Ed. by B. R. Pridham, pp. 154–174. London – Sydney: Croom Helm.
Dresch P. 1989. Tribes, Government, and History in Yemen. Oxford: Clarendon Press.
Dresch P. 2016. The Rules of Barat. Tribal Documents from Yemen: Texts and Translation. Paris: Centre français d’archéologie et de sciences sociales.
Dunbar R. I. M. 1988. Primate Social Systems. London: Crook Helm.
Dunbar R. I. M. 1994. Ecological Constraints on Group Size in Baboons. Animal
Societies: Individuals, Interactions and Social Organization / Ed. by P. Jarman,
and
A. Rossiter, pp. 221–236. Oxford: Blackwell.
Earle T. K. 1987. Chiefdoms in Archaeological and Ethnohistorical Perspective. Annual Review of Anthropology 16: 279–308.
Ehrenreich P. 1887. Über die Botocudos der Brasilianischen Provinzen Espiritu Santo und Minas Geraes. Zeitschrift für Ethnologie 19: 1–46.
Eibl-Eibesfeldt I., and Sutterlin C. 1992. Im Banne Der Angst. Muenchen: R. Piper GmbH.
Elkin A. P. 1954. The Australian Aborigines: How to Understand Them. Sydney. Angus and Robertson.
Embree J. F. 1939. Suye Mura: A Japanese Village. The University of Chicago Publications in Anthropology, Ethnological Series. Chicago: University of Chicago Press.
Ember M., and Ember C. R. 1999. Anthropology. 9th ed. Upper Saddle River, NJ: Prentice Hall.
Emery Thompson M. 2019. How Can Non-Human Primates Inform Evolutionary Perspectives on Female-Biased Kinship in Humans? Philosophical Transactions of the Royal Society B 374(1780): 20180074.
Engelhart M. 1998. Extending the Tracts: A Cross-Reductionist Approach to Australian Aboriginal Male Initiation Rites. Acta Universitatis Stockholmiensis, Stockholm Studies in Comparative Religion 34. Stockholm: Almqvist and Wiksel.
Estrada A. 1984. Male-Infant Interactions Among Free-Ranging Stumptail Macaques. Primate Paternalism / Ed. by D. M. Taub, pp. 56–87. New York: Van Nostrand Reinhold.
Fedosova V. N. 1994. The Development of Modern Palaeodemography (Palaeoecological Aspects of Data Analysis). Rossijskaja arheologija 3: 71–82. In Russian (Федосова В. Н. Развитие современной палеодемографии (палеоэкологические аспекты анализа фактических данных). Российская археология 3: 71–82).
Frantsouzoff S. A. 1995. The Inscriptions from the Temples of Dhat Himyam at Raybun. Proceedings of the Seminar for Arabian Studies 25: 15–27.
Frantsouzoff S. A. 1997. Regulation of Conjugal Relations in Ancient Raybun. Proceedings of the Seminar for Arabian Studies 27: 47–62.
Frantsouzoff S. A. 2000. The Society of Raybûn. Alternatives
of Social Evolution / Ed. by N.
N. Kradin, A. V. Korotayev, V. de Munck, V. A. Lynsha, D. M. Bonda-
renko, and P. Wason, pp. 258–265. Vladivostok: Dal'nauka.
Franz C. 1999. Allogrooming Behavior and Grooming Site Preferences in Captive Bonobos (Pan Paniscus): Association With Female Dominance. Int. J. Primatol. 20: 525–546.
Fried M. H. 1967. The Evolution of Political Society; An Essay in Political Anthropology. New York: Random House.
Friedman J., and Rowlands M. J. 1977. Notes Towards an Epigenetic Model of ‘Civilization’. The Evolution of Social Systems / Ed. by J. Friedman, and M. J. Rowlands, pp. 201–276. London: Duckworth.
Geertz H., and Geertz C. 1975. Kinship in Bali. Chicago: Chicago University Press.
Goodale J. C. 1962. Marriage Contracts Among the Tiwi. Ethnology 1: 452–466.
Goodale J. C. 1971. Tiwi Wives. Seattle – London: University of Washington Press.
Gore M. S. 1965. The Traditional Indian Family. Comparative Family Systems / Ed. by M. F. Nimkoff, pp. 209–231. Boston: Houghton-Mifflin.
Gould R. A. 1969. Yiwara: Foragers of the Australian Desert. New York: Scribner.
Gould R. A. 1981. Comparative Ecology of Food-Sharing in Australia and Northwest California. Omnivorous Primates / Ed. by S. O. Harding, and G. Teleki, pp. 422–454. New York.
Gould R. A. 1991. Arid-Land Foraging as Seen from Australia: Adaptive Models and Behavioral Realities. Oceania 62(1): 12–33.
Grinin L. E., and Korotayev A. V. 2009. The Epoch of the Initial Politogenesis. Social Evolution & History 8(1): 52–91.
Grinin L. E., and Korotayev A. V. 2011. Chiefdoms and Their Analogues: Alternatives of Social Evolution at the Societal Level of Medium Cultural Complexity. Social Evolution and History 10(1): 276–335.
Grinin L. E., and Korotayev A. V. 2017. Chiefdoms and Their Analogues: Alternatives of Social Evolution at the Societal Level of Medium Cultural Complexity. Chiefdoms. Yesterday and Today / Ed. by R. L. Carneiro, L. E. Grinin, and A. V. Korotayev, pp. 63–108. New York: Eliot Werner Press.
Grinin L. E., Markov A. V., and Korotayev A. V. 2009. Aromorphoses in Biological and Social Evolution: Some General Rules for Biological and Social Forms of Macroevolution. Social Evolution and History 8(2): 6–50.
Grinin L. E., Markov A. V., and Korotayev A. V. 2011. Biological and Social Aromorphoses: A Comparison between Two Forms of Macroevolution. Evolution: Cosmic, Biological, and Social / Ed by L. E. Grinin, R. L. Carneiro, A. V. Korotayev, and F. Spier, pp. 162–211. Volgograd: Uchitel.
Guenther M. 1986. The Nharo Bushmen of Botswana: Tradition and Social Change. Hamburg: Helmut Buske Verlag (Quellen zur Khoisan Forschung 3).
Gurevich A. Ya. 1972. History and the Saga. Moscow: Nauka. In Russian (Гуревич А. Ю. История и сага. М.: Наука).
Gusinde M. 1956. Die Twiden: Pygmäen und Pygmöide im Tropischen Afrika. Vienna – Stuttgart: Wilhelm Braumüller.
Harcourt A. H. 1978. Strategies of Emigration and Transfer by Primates, with Particular Reference to Gorillas. Z. Thierpsychol. 48: 401–420.
Hart C. W. M. 1930. The Tiwi of Melville and Bathurst Islands. Oceania 1: 167–290.
Hausfater G., Altmann J., and Altmann
S. 1982. Long-term Consistency of Dominance Relations Among Female Baboons (Papio cynocephalus). Science 217:
752–755.
Haviland W. A. 1991. Anthropology. 6th ed. Fort Worth, TX: Holt, Rinehart and Winston.
Heinz H. J. 1972. Territoriality Among the Bushmen in General and the !Ko in Particular. Anthropos 67(4): 405–416.
Hewitt R. L. 1986. Structure, Meaning and Ritual in the Narration of the Southern San. Hamburg: Helmut Buske Verlag (Quellen zur Khoisan Forschung 2).
Hiatt L. 1959. Social Control in Central Arnhem Land. South Pacific 10: 182–192.
Hinde R. 1983. Primate Social Relationships: An Integrated Approach. Oxford: Blackwell.
Hohmann G., and Fruth B. 1993. Field Observations on Meat Sharing Among Bonobos (Pan paniscus). Folia Primatol. 60: 225–229.
van Hooff J. A., and van Schaik C. P. 1994. Male Bonds: Affiliative Relationships Among Nonhuman Primate Males. Behavior 130: 309–337.
Hrdy S. B., and Burkart J. M. 2020. The Emergence of Emotionally Modern Humans: Implications for Language and Learning. Philosophical Transactions of the Royal Society B 375(1803): 20190499.
Ibn Al-Mu`tazz. 1961. Diwân. Bayru:t: Dâr al-Thaqâfah.
Ihobe H. 1992. Male-Male Relationships Among Wild Bonobos (Pan paniscus) at Wamba, Republic of Zaire. Primates 33: 163–179.
Ishii-Kuntz M., and Lee G. R. 1987. Status of the Elderly: An Extension of the Theory. Journal of Marriage and Family 49: 413–420.
Jin F. F. 1996. Tropical Ocean – Atmosphere Interaction: The Pacific Cold Tongue and El Niño-Southern Oscillation. Science 274: 76–78.
Judge P. G. 1991. Dyadic and Triadic Reconciliation in Pigtail Macaques (Macaca nemestrina). Am. J. Primatol. 23: 225–237.
Goodall J. 1986. The Chimpanzees of Gombe: Patterns of Behavior. Cambridge: Harvard University Press.
Kabo V. R. 1986. Primitive Preagricultural Community. Moscow: Nauka. In Russian (Кабо В. Р. Первобытная доземледельческая община. М.: Наука).
Kalbitzer U., Bergstrom M. L., Carnegie S. D., Wikberg E. C., Kawamura S., Campos F. A. et al. 2017. Female Sociality and Sexual Conflict Shape Offspring Survival in a Neotropical Primate. Proceedings of the National Academy of Sciences 114(8): 1892–1897.
Kano T. 1992. The Last Ape: Pygmy Chimpanzee Behavior and Ecology. Stanford, CA: Stanford University Press.
Kappeler P. M. 2019. A Framework for Studying Social Complexity. Behavioral Ecology and Sociobiology 73(1): 13.
Keen I. 1994. Knowledge and Secrecy in an Aboriginal Religion. Oxford: Clarendon Press.
Korotayev A. 1995a. Ancient Yemen. Some General Trends of Evolution of the Sabaic Language and the Sabaean Culture. Oxford – New York: Oxford University Press.
Korotayev A. V. 1995b. Mountains and Democracy: An Introduction. Alternative Pathways to Early State / Ed. by N. N. Kradin, and V. A. Lynsha, pp. 60–74. Vladivostok: Dal'nauka.
Korotayev A. V. 1995c. Some Problems of Social Evolution of Archaic (and not only Archaic) Societies. Vostok (5): 211–220. In Russian (Коротаев А. В. Некоторые проблемы социальной эволюции архаических (и не только архаических) обществ. Восток (5): 211–220.
Korotayev A. V. 1995d. Apologia for Tribalism: Tribe as a Form of Socio-Political Organization of Complex Non-Primitive Societies. Sotsiologicheskij zhurnal 4: 68–86. In Russian (Коротаев А. В. Апология трайбализма: племя как форма социально-политической организации сложных непервобытных обществ. Социологический журнал 4: 68–86).
Korotayev A. V. 1996a. Pre-Islamic Yemen. Sociopolitical Organization of the Sabaean Cultural Area in the 2nd and 3rd Centuries A.D. Wiesbaden: Harrassowitz Verlag.
Korotayev A. V. 1996b. Two Socio-Ecological Crises and Genesis of Tribal Organization in the Yemeni North-East. Vostok 6: 18–28. In Russian (Коротаев А. В. Два социально-экологических кризиса и генезис племенной организации на северо-востоке Йемена. Восток 6: 18–28).
Korotayev A. V. 1996c. From Chiefdom to Tribe? Some Trends of Political Evolution of North-East Yemen in the Last Two Millennia. Etnograficheskoe obozrenie 2: 81–91. In Russian (Коротаев А. В. От вождества к племени? Некоторые тенденции эволюции политических систем северо-восточного Йемена за последние две тысячи лет. Этнографическое обозрение 2: 81–91).
Korotayev A. V. 1997. Sabaean Studies: Some General Trends and Factors of Evolution of the Sabaean Civilization. Мoscow: Vostochnaja literatura. In Russian (Коротаев А. В. Сабейские этюды: некоторые общие тенденции и факторы эволюции сабейской цивилизации. М.: Восточная литература).
Korotayev A. V. 1998. Chiefdoms and Tribes of the Land of Ha:shid and Baki:l. Moscow: IV RAN. In Russian (Коротаев А. В. Вождества и племена страны Хашид и Бакил. M.: ИВ РАН).
Korotayev A. V. 1999. The Correlation between the Kinship Terms and Social System Types: A Quantitative Cross-Cultural Comparison. Kinship Algebra 3: 117–147. In Russian (Коротаев А. В. О соотношении систем терминов родства и типов социальных систем: опыт количественного кросс-культурного анализа. Алгебра родства 3: 117–147).
Korotayev A. V. 2000. The Chiefdom: Precursor of the Tribe? (Some Trends of the Evolution of the Political Systems of the North-East Yemen in the 1st and 2nd Millennia A.D.). Alternatives of Social Evolution / Ed. by N. N. Kradin, A. V. Korotayev, V. de Munck, V. A. Lynsha, D. M. Bondarenko, and P. Wason, pp. 242–257. Vladivostok: Dal'nauka.
Korotayev A. 2003a. Christianity and Democracy: A Cross-Cultural Study (Afterthoughts). World Cultures 13(2): 195–212.
Korotayev A. V. 2003b. Penetration of Nomads to the Arabian South and Formation of Tribal Organization among the North-East Yemen Agricultural Population. Nomadic Pathways in Social Evolution / Ed. by N. N. Kradin, D. M. Bondarenko, and Th. J. Barfield, pp. 114–122. Moscow: Institute for African Studies.
Korotayev A. 2003c. Unilineal Descent Groups and Deep Christianization: A Cross-Cultural Comparison. Cross-Cultural Research 37(1): 132–156.
Korotayev A. 2004. World Religions and Social Evolution of the Old World Oikumene Civilizations: A Cross-Cultural Perspective. Lewiston, NY: The Edwin Mellen Press.
Korotayev A. V. 2006. Social History of Yemen. Moscow: KomKniga/URSS. In Russian (Коротаев А. В. Социальная история Йемена. М.: КомКнига/URSS).
Korotayev A. V. 2020. Evolution of Sociopolitical Institutions in North-East Yemen (The 1st Millennium BCE – The 2nd Millennium CE). The Evolution of Social Institutions. Interdisciplinary Perspectives / Ed. by D. M. Bondarenko, S. A. Kowalewski, and D. B. Small, pp. 161–184. Cham: Springer. DOI: 10.1007/978-3-030-51437-2_7.
Korotayev A. V., and Bondarenko D. M. 2000. Polygyny and Democracy: A Cross-Cultural Comparison. Cross-Cultural Research 34(2): 190–208.
Korotayev A., Borinskaya S., Starostin G., and Meshcherina K. 2019. Evolution of Eurasian and African Family Systems, Cross-Cultural Research, Comparative Linguistics, and Deep History. Social Evolution & History 18(2): 286–312. DOI: 10.30884/seh/2019.02.15.
Kottak C. Ph. 2000. Anthropology. The Exploration of Human Diversity. 8th ed. Boston, MA: McGraw-Hill.
Kozintsev A. G. 1980. The Transition to Farming and the Ecology of Man. Early Farmers: Ethnographic Essays / Ed. by N. A. Butinov, and A. M Reshetov, pp. 6–33. Leningrad: Nauka. In Russian (Козинцев А. Г. Переход к земледелию и экология человек. Ранние земледельцы: Этнографические очерки / Ред. Н. А. Бутинов, А. М. Решетов, с. 6–33. Л.: Наука).
Kradin N. N. 2006. Cultural Complexity of Pastoral Nomads. World Cultures 15(2): 171–189.
Kradin N. N. 2008. Structure of Power in Nomadic Empires of Inner Asia: Anthropological Approach. Hierarchy and Power in the History of Civilizations: Ancient and Medieval Cultures / Ed. by L. E. Grinin, D. D. Belyaev, and A. V. Korotayev, pp. 98–124. Moscow: URSS.
Kradin N. N. 2011a. Heterarchy and Hierarchy among the Ancient Mongolian Nomads. Social Evolution and History 10(1): 187–214.
Kradin N. N. 2011b. Stateless Empire: The Structure of the Xiongnu Nomadic Super-Complex Chiefdom. Xiongnu Archaeology: Multidisciplinary Perspectives of the First Steppe Empire in Inner Asia / Ed. by U. Brosseder, and B. K. Miller, pp. 77–96. Bonn: Rheinische Friedrich-Wilhelms-Universität.
Kradin N. N. 2014. Nomads of Inner Asia in Transition. Moscow: Krasand.
Kradin N. N. 2018. Historical Dynamics and Succession of Inner Asian Nomadic Empires. Chronica: Annual of the Institute of History University of Szeged 18: 191–197.
Kradin N. N. 2019. Social Complexity, Inner Asia, and Pastoral Nomadism. Social Evolution and History 18(2): 3–34.
Kradin N. N. 2020. Heterarchy and
Hierarchy in Eurasian Steppes. The
Evolution of Social Institutions.
Interdisciplinary Perspectives / Ed. by D. M. Bondarenko, S. A. Ko-
walewski, and D. B. Small, pp. 185–204.
Cham: Springer. DOI: 10.1007/978-3-030-51437-2_7.
Kuester J., and Paul A. 1986. Male–Infant Relationships in Semi-Free Ranging Barbary Macaques (Macaca sylvanus) of Affenberg Salem/FRG: Testing the ‘Male Care’ Hypothesis. Am. J. Primatol. 10: 315–327.
Kuester J., and Paul A. 1992. Influence of Male Competition and Female–Male Choice on Male Mating Success in Barbary Macaques (Macaca sylvanus). Behavior 120: 192–217.
Kummer H. 1971. Primate Societies. Chicago: Aldine.
Kuroda S. 1984. Interactions over Food Among Pygmy Chimpanzees. The Pygmy Chimpanzee: Evolutionary Biology and Behavior / Ed. by R. L. Susman, pp. 301–324. New York: Plenum.
Lamilami L. 1974. Lamilami Speaks: The Cry Went On. Sydney: Ure Smith.
Lancaster J. B. 1997. The Evolutionary History of Human Parental Investment in Relation to Population Growth and Social Stratification. Feminism and Evolutionary Biology / Ed. by P. A. Gowaty, pp. 466–488. New York: Chapman and Hall.
Layton R. 1986. Uluru: An Aboriginal History of Ayers Rock. Canberra: Australian Institute for Aboriginal Studies.
Lee Gary R. 1984. Status of the Elderly: Economic and Familial Antecedents. Journal of Marriage and Family 46: 267–275.
Lee Gary R., and Kezis M. 1979. Family Structure and the Status of the Elderly: A Preliminary Study. Journal of Comparative Family Studies 10: 429–443.
Lee Kwang-Kyu. 1975. Kinship System in Korea. 2 vols. New Haven: Yale University Press.
Lee R. B. 1972. The !Kung Bushmen of Botswana. Hunters and Gatherers Today / Ed. by M. G. Bicchieri, pp. 327–368. New York: Holt, Rinehart and Winston.
Lee R. B. 1979. The !Kung San: Men, Women and Work in a Foraging Society. Cambridge: Cambridge University Press.
Lee R. B., and Devore I. 1968. Problems in the Study of Hunters and Gatherers. Man the Hunter / Ed. by R. B. Lee, and I. DeVore, pp. 3–20. Chicago: Aldine.
Leigh J. H., and Noble J. C. 1969. Vegetation Resources. Arid Lands of Australia / Ed. by R. O. Slayter, and R. A. Perry, pp. 73–92. Canberra: Australian National University.
Leistner O. A. 1979. Southern Africa. Arid Land Ecosystems / Ed. by D. W. Goodall, vol. 1, pp. 109–143. Cambridge: Cambridge University Press.
Liberman K. 1985. Understanding Interaction in Central Australia. Boston: Routledge and Kegan Paul.
Livi-Bacci M. 2012. A Concise History of World Population. 5th ed. Malden, MA: Wiley-Blackwell.
Makal M. 1954. A Village in Anatolia. London: Vallentine, Mitchell and Co.
Manguette M. L., Robbins A. M., Breuer T., Stokes E. J., Parnell R. J., and Robbins M. M. 2019. Intersexual Conflict Influences Female Reproductive Success in a Female-Dispersing Primate. Behavioral Ecology and Sociobiology 73(9): 118.
Marshall L. 1965. The !Kung Bushmen of the Kalahari Desert. Peoples of Africa / Ed. by J. L. Gibbs, Jr., pp. 241–278. New York: Holt, Rinehart and Winston.
Marshall L. 1976. The !Kung of Nyae Nyae. Cambridge, MA: Harvard University Press.
Matsumura S. 1996. Post-Conflict Affiliative Contacts Between Former Opponents Among Wild Moor Macaques (Macaca maurus). Amer. J. Primatol. 38: 211–219.
Matsumura S. 1999. The Evolution of ‘Egalitarian’ And ‘Despotic’ Social Systems Among Macaques. Primates 43: 23–31.
Matthew B. et al. 1999. Horse Sickness and ENSO in South Africa. Nature 397: 574.
Mcnett C. W. 1973. Factor Analysis of a Cross-Cultural Sample. Behavior Science Notes 8: 233–257.
Meggitt M. J. 1962. Desert People. Sydney: Angus and Robertson.
Mel'jantsev V. A. 1996. East and West in the 2nd Millennium: Economics, History, and Modernity. Moscow: Izdatel'stvo Moskovskogo universiteta. In Russian (Мельянцев В. А. Восток и Запад во втором тысячелетии: экономика, история и современность. М.: Издательство Московского университета).
Meshkov K. Yu. 1982. The Philippines. Small Nations of Indonezia, Malaysia, and the Philippines / Ed. by N. N. Cheboksarov, and A. I. Kuznetsov, pp. 175–226. Mosсow: Nauka. In Russian (Мешков К. Ю. Филиппины. Малые народы Индонезии, Малазии и Филиппин / Ред. Н. Н. Чебоксаров, А. И. Кузнецов, с. 175–226. М.: Наука).
Mitchell C. L., Boinski S., van Schaik C. P. 1991. Competitive Regimes and Female Bonding in Two Species of Squirrel Monkeys (Saimiri oerstedi and S. sciereus). Behavioral Ecology and Sociobiology 28: 55–60.
Moore
J. 1996. Savanna Chimpanzees, Referential Models and the Last
Common Ancestor. Great Apes Societies / Ed. by W. C. McGrew, L. Marchant,
and T. Ni-
shida, pp. 275–292. Cambridge: Cambridge University Press.
Moscovice L. R., Sueur C., and Aureli F. 2020. How Socio-Ecological Factors Influence the Differentiation of Social Relationships: An Integrated Conceptual Framework. Biology Letters 16(9): 20200384.
Mountford Ch. P. 1958. The Tiwi: Their Art, Myth, and Ceremony. London: Phoenix House.
de Munck V., and
Korotayev A. 1999. Sexual Equality and Romantic Love:
A Reanalysis of Rosenblatt's Study on the Function of Romantic Love. Cross-Cultural
Research. The Journal of Comparative Social
Science 33: 265–277.
Murdock G. P. 1967. Ethnographic Atlas: A Summary. Pittsburgh: The University of Pittsburgh Press.
Murdock G. P. 1981. Atlas of World Cultures. Pittsburgh: The University of Pittsburgh Press.
Murdock G. P., and Provost C. 1971. Measurement of Cultural Complexity. Ethnology 12: 379–392.
Murdock G. P., and Provost C. 1985. Measurement of Cultural Complexity. World Cultures 1(1): stds6.dat.
Murdock G. P., and White D. R. 1969. Standard Cross-Cultural Sample. Ethnology 8: 329–369.
Murdock G. P., Textor R., Barry H. (III), and White D. R. 1986. Ethnographic Atlas. World Cultures 2(4) (the first computer version).
Murdock G. P., Textor R., Barry H. (III), and White D. R. 1990. Ethnographic Atlas. World Cultures 6(3) (the second computer version).
Murdock G. P.,
Textor R., Barry H. (III), White D. R., Gray J. P., and
Divale W. 1999. Ethnographic Atlas. World Cultures 10(1): 24–136, ea01–09.sav (the third computer version).
Muthukrishna M., Doebeli M., Chudek M., and Henrich J. 2018. The Cultural Brain Hypothesis: How Culture Drives Brain Expansion, Sociality, and Life History. PLoS Computational Biology 14(11): e1006504.
Myers
F. R. 1980. The Cultural Basis of Politics in Pintupi Life. Mankind 12(3):
197–214.
Nikitin N. I. 1987. On
the Formational Nature of Early Cossack Communities. An Introduction. Feudalism
in Russia / Ed. by V. L. Yanin, pp. 236–245. Moscow: Nauka. In Russian (Никитин Н. И. О
формационной природе ранних казачьих сообществ (к постановке вопроса). Феодализм в России / Ред. В. Л. Янин,
с. 236–245. М.: Наука).
Nimkoff M. F., and Middleton R. 1960. Types of Family and Types of Economy. American Journal of Sociology 66: 215–225.
Nimuendajú C. 1946. Social Organization and Beliefs of the Botocudo of Eastern Brazil. Southwestern Journal of Anthropology 2(1): 93–115.
Nishida
T., and Hiraiwa-Hasegawa M. 1987. Chimpanzees and Bonobos: Cooperative
Relationships Among Males. Primate Societies / Ed. by B. Smuts, D. L. Cheney,
R. M. Seyfarth, R. W. Wrangham, and T. T. Struhsaker, pp. 165–177. Chicago, IL:
University of Chicago Press.
Noe P. 1986. Lasting Alliances Among Adult Male Savanna Baboons. Primate Ontogeny, Cognition and Social Behavior / Ed. by J. G. Lee, pp. 381–392. New York – Cambridge: Cambridge University Press.
van Noordwijk M. A., and van Schaik C. P. 1987. Competition among Female Long-Tailed Macaques, Macaca fascicularis. Animal Behavior 35: 577–589.
Norbeck E. 1954. Takashima: A Japanese Fishing Community. Salt Lake City, UT: University of Utah Press.
Obermeyer
G. J. 1982. Le formation de l'imamat et de l'état au Yémen: Islam
et culture politique. La péninsule Arabique d'aujourd'hui. T. II. Etudes
par pays / Ed. by
P. Bonnenfant, pp. 31–48. Paris: CNRS.
Ogawa H. 1995. Bridging Behavior and Other Affiliative Interactions Among Tibetan Macaques (Macaca thibetana). Intern. J. Primatol. 16(5): 707–729.
Ogawa H. 2020. Bridging Behavior and Male-Infant Interactions in Macaca Thibetana and M. Assamensis: Insight into the Evolution of Social Behavior in the Sinica Species-Group of Macaques. The Behavioral Ecology of the Tibetan Macaque / Ed. by Jin-Hua Li, Lixing Sun, and P. Kappeler, pp. 189–203. Cham: Springer.
Olgeirsson E. 1957. The Past of the Icelandic People. Moscow: Politizdat. In Russian (Ольгейрссон Э. Из прошлого исландского народа. М.: Политиздат).
Osmond M. W. 1969. A Cross-Cultural Analysis of Family Organization. Journal of Marriage and Family 31: 302–310.
Ostner
J., and Schuelke O. 2018. Linking Sociality to
Fitness in Primates: A Call For Mechanisms. Vol. 50. Advances in the Study
of Behavior / Ed. by M. Naguib,
L. Barrett, S. D. Healy, J. Podos, L. W.
Simmons, and M. Zuk, pp. 127–175.
Owens M., and Owens D. 1985. Cry of the Kalahari. Boston: Houghton Mifflin.
Palombit R. A., Seyfarth R. M., and Cheney D. L. 1997. The Adaptive Value of ‘Friendship’ To Female Baboons: Experimental and Observational Evidence. Anim. Behav. 54: 599–614.
Pasternak B. 1976. Introduction to Kinship and Social Organization. Englewood Cliffs, NJ: Prentice Hall.
Pasternak B., Ember M., and Ember C. 1997. Sex, Gender, and Kinship: A Cross-Cultural Perspective. Upper Saddle River, NJ: Prentice Hall.
Paul A., and Kuester J. 1987. Dominance, Kinship and Reproductive Value in Female Barbary Macaques (Macaca sylvanus) at Affenberg Salem. Behav. Ecol Sociobiol. 21: 323–331.
Petit O., Abegg C., and Thierry B. 1997. A Comparative Study of Aggression and Conciliation in Three Cercopithecine Monkeys (Macaca fuscata, Macaca nigra, and Papio papio). Behavior 134: 415–432.
Petit O., and Thierry B. 1994a. Aggressive and Peaceful Interventions in Conflicts in Tonkean Macaques. Animal Behavior 48: 89–95.
Petit O., and Thierry B. 1994b. Reconciliation in a Group of Black Macaques (Macaca Nigra). Dodo. Journal of Wildlife Preservation Trusts 30: 89–95.
Picaut J., Ioualalen M., Menkes C., Delcroix T., McPhaden M. J. 1996. Mechanism of the Zonal Displacement of the Pacific Warm Pool: Implications for ENSO. Science 274: 1486–1489.
Piddington R. 1932–1933. Karadjeri Initiation. Oceania 3: 46–87.
Porteus S. D. 1931. The Psychology of the Primitive People. New York: Longmans, Green and Co.
Precourt W. 1975. Initiation Ceremonies and Secret Societies as Educational Institutions. Cross-Cultural Perspectives on Learning / Ed. by R. Brislin, W. Lonner, and S. Boohner, pp. 231–250. Beverly Hills, CA: Sage.
Preuschoft S., Paul A., and Kuester J. 1998. Dominance Styles of Female and Male Barbary Macaques (Macaca fuscata). Behavior 135: 731–755.
Rodseth L., Wrangham R. W., Harrigan A. M., and Smuts B. B. 1991. The Human Community as a Primate Society. Current Anthropology 32: 221–254.
Rosenblatt P. 1967. Marital Residence and the Functions of Romantic Love. Ethnology 6: 471–480.
Ross M. H. 1983. Political Decision Making and Conflict: Additional Cross-Cultural Codes and Scales. Ethnology 22: 169–192.
Ross M. H. 1986. Political Decision Making and Conflict: Additional Cross-Cultural Codes and Scales. World Cultures 2(2): stds30.dat.
Rozner I. G. 1970. Antifeudal States in Russia and Ukraine in the 16th – 18th Centuries. Voprosy istorii (8): 42–56. In Russian (Рознер И. Г. Антифеодальные государственные образования в России и на Украине в XVI–XVIII вв. Вопросы истории 8: 42–56).
Sahlins M. D. 1960. Evolution: Specific and General.
Evolution and Culture / Ed. by
M. D. Sahlins, and E. R. Service, pp. 12–44. Ann Arbor, MI: University of Michigan
Press.
Sahlins M. D. 1972. Stone Age Economics. London: Aldine.
Saito C. 1996. Dominance and Feeding Success in Female Japanese Macaques, Macaca fuscata: Effects of Food Patch Size and Inter-Patch Distance. Animal Behavior 51: 967–980.
Sayfarth R. M. 1978. Social Relationships Among Adult Male and Female Baboons. II. Behavior Throughout the Female Reproductive Cycle. Behavior 64: 227–247.
SCCS. 1999. Standard Cross-Cultural Sample Variables / Ed. by W. Divale. World Cultures 10(1): stds01–83.sav. URL: https://doi.org/10.13140/RG.2.2.18058.29120.
van Schaik C. P. 1989. The Ecology of Social Relationships Amongst Female Primates. Comparative Socioecology. The Behavioral Ecology of Humans and Other Mammals / Ed. by V. Standen, and R. A. Foley, pp. 195–218. Oxford: Oxford University Press.
van Schaik C. P., and van Hooff J. A. R. A. M. 1983. On the Ultimate Causes of Primate Social Systems. Behaviour 85(1–2): 91–117.
van Schaik C. P., van Noordwijk M., and Nunn C. L. 1999. Sex and Social Evolution in Primates. Comparative Primate Socioecology / Ed. by P. C. Lee, pp. 204–240. Cambridge: Cambridge University Press.
Schebesta P. 1933. Among Congo Pygmies. London: Hutchinson.
Schebesta P. 1948. Die Bambuti-Pygmäen vom Ituri. Brussels: Institut Royal Colonial Belge.
Schlegel A., and Barry H. III. 1980. The Evolutionary Significance of Adolescent Initiation Ceremonies. American Ethnologist 7: 696–715.
Service E. R. 1971 [1962]. Primitive Social Organization. An Evolutionary Perspective. 2nd ed., New York, NY: Random House.
Sicotte P. 1993. Inter-group Interactions and Female Transfer in Mountain Gorillas: Influence of Group Composition on Male Behavior. Amer. J. Primatol. 30: 21–36.
Sicotte P. 1995. Interpositions in Conflicts Between Males in Bimale Groups of Mountain Gorillas. Folia Primatol. 65: 14–24.
Sigg H. 1980. Differentiation of Female Positions in Hamadryas One-Male Units. Z. Tierpsychol 53: 265–302.
Silberbauer G. B. 1972. The G/wi Bushmen. Hunters and Gatherers Today / Ed. by M. G. Bicchieri, pp. 271–326. New York: Holt, Rinehart and Winston.
Silberbauer G. B. 1981. Hunter and the Habitat in the Central Kalahari Desert. Cambridge: Cambridge University Press.
Silk J. B. 1982. Altruism Among Female Macaca Radiata: Explanations and Analysis of Patterns of Grooming and Coalition Formation. Behaviour 79(2–4): 162–188.
Silk J., Cheney D. L., and Seyfarth R. M. 1996. The Form and Function of Post-Conflict Interactions Between Female Baboons. Animal Behavior 52: 259–268.
Small D. B. 2019. Ancient Greece: Social Structure and Evolution. Cambridge: Cambridge University Press.
Small D. B. 2020. Institutional Evolution of Ancient Greece. The Evolution of Social Institutions. Interdisciplinary Perspectives / Ed. by D. M. Bondarenko, S. A. Kowalewski, and D. B. Small, pp. 255–282. Cham: Springer. DOI: 10.1007/978-3-030-51437-2_11.
Spencer B., and Gillen F. J. 1899. The Native Tribes of Central Australia. London: Macmillan.
Spencer B., and Gillen F. J. 1927. The Arunta. London: Macmillan.
Steblin-Kamenskij M. I. 1984. The World of the Saga. Leningrad: Nauka. In Russian (Стеблин-Каменский М. И. Мир саги. Л.: Наука).
Sterck E. H., and Watts D. W., and van Schaik C. P. 1997. The Evolution of Female Social Relationships in Nonhuman Primates. Behav. Ecol. Sociobiol. 41: 291–309.
Storey R. 1985. An Estimate of Mortality in a Pre-Columbian Urban Population. American Anthropologist 87: 515–535.
Strehlow T. G. H. 1947. Aranda Traditions. Melbourne: Melbourne University Press.
Strehlow T. G. H. 1965. Culture, Social Structure
and Environment in Aboriginal Central Australia. Aboriginal Man in Australia
/ Ed. by R. M. Berndt, and
C. H. Berndt, pp. 121–145. Sydney: Angus and Robertson.
Street S. E., Navarrete A. F., Reader S. M., and Laland K. N. 2017. Coevolution of Cultural Intelligence, Extended Life History, Sociality, and Brain Size in Primates. Proceedings of the National Academy of Sciences 114(30): 7908–7914.
Strier K. B. 1999. Why is Female Kin Bonding So
Rare? Comparative Sociality of Neotropical Primates. Comparative Primate Socioecology
/ Ed. by P. C. Lee,
pp. 300–319. Cambridge: Cambridge University Press.
Sugawara K. 1988. Ethological Study of the Social Behavior of Hybrid Baboons Between Papio Anubis and P. hamadryas in Free-Ranging Groups. Primates 29: 429–448.
Sugiyama Y. 1989. Population Dynamics of Chimpanzees at Bossou, Guinea. Understanding Chimpanzees / Ed. by P. Heltne, and L. Marquardt, pp. 134–145. Cambridge, MA: Harvard University Press.
Susman R. L. 1987. Pygmy Chimpanzees and Common Chimpanzees: Models for the Behavioral Ecology of the Earliest Hominids. The Evolution of Human Behavior: Primate Models / Ed. by W. G. Kinzey, pp. 72–86. Albany, NY: State University of New York Press.
Takahata Y., Sprague D. S., Suzuki S., and Okayasu N. 1994. Female Competition, Co-Existance, and the Mating Structure of Wild Japanese Macaques on Yakushima Island, Japan. Animal Societies: Individuals, Interactions and Organisation / Ed. by P. J. Jarman, and A. Rossiter, pp. 163–179. Kyoto: Kyoto University Press.
Taub
D. M. 1980. Female Choice and Mating Strategies among Wild Barbary
Macaques (Macaca sylvanus). The Macaques: Studies in Ecology, Behavior and
Evoluti-
on / Ed. by D. G. Lindberg, pp. 287–344. New York: Van Nostrand Reinhold.
Terry M. 1928. War of the Waramullas. Adelaide: Rigby.
Thierry B. 1985. Patterns of Agonistic Interactions in Three Species of Macaque (Macaca mulatta, M. fascicularis, M. tonkeana). Aggressive Behavior 11: 223–233.
Thierry B. 1988. A Comparative Study of Aggression and Response to Aggression. Three Species of Macaques. Primate Ontogeny, Cognition, and Social Behavior / Ed. by J. Else, and P. C. Lee, pp. 307–313. Cambridge: Cambridge University Press.
Thierry B. 1990. Feedback Loop Between Kinship and Dominance: The Macaque Model. J. Theor. Biol. 145: 511–521.
Thierry B. 1999. Covariation of Conflict Management Patterns Across Macaque Species. Natural Conflict Resolution / Ed. by F. Aureli, and F. de Waal. Berkeley: University of California Press.
Thierry B., Gauthier C., and Peignot P. 1990. Social Grooming in Tonkean Macaques (Macaca tonkeana). Int. J. Primatol. 11: 357–375.
Thomas E. M. 1959. The Harmless People. New York: Alfred A. Knopf.
Threlkeld L. E. 1974. Australian Reminiscences and Papers of L. E. Threlkeld, Missionary to the Aborigines, 1824–1859. 2 vols. Canberra: Australian Institute of Aboriginal Studies.
Tindale N. B. 1935–1936. Initiation Among the Pitjandjara Natives of the Mann and Tomkinson Ranges in South Australia. Oceania 6: 200–224.
Tindale N. B. 1972. Pitjantjatjara. Hunters and Gatherers Today / Ed. by M. G. Bicchieri, pp. 98–134. New York: Holt, Rinehart and Winston.
Tolstov S. P. 1962. Peoples of the Caucasus. Vol. 2. Moscow: Izdatel'stvo Akademii Nauk SSSR. In Russian (Толстов С. П. Народы Кавказа. Том 2. М.: Издательство Академии Наук СССР).
Tonkinson R. 1974. Aboriginal Victors of the Desert Crusade. Menlo Park, CA: Benjamin.
Townsend J. B. 1985. The
Autonomous Village and the Development of Chiefdoms. Development and Decline.
The Evolution of Sociopolitical Organization / Ed. by
H. J. M. Claessen, P. van de Velde, and M. E. Smith, pp. 141–155. South Hadley,
MA: Bergin & Garvey.
Turnbull C. M. 1961. The Forest People. New York, NY: Simon and Schuster.
Turnbull C. M. 1965a. Wayward Servants. Garden City, NY: Natural History Press.
Turnbull C. M. 1965b. The Mbuti Pygmies. Anthropological Papers of the American Museum of Natural History 50(1): 1–282.
Tutin C. E., Mcgrew W. C., and Baldwin P. J. 1983. Social Organization of Savanna-dwelling Chimpanzees, Pan troglodytes verus, at Mt. Assirik, Senegal. Primates 24(2): 154–173.
Vasil'jev L. S. 1980. The Formation of Political Administration (From a Local Group of Hunter-Gatherers to a Chiefdom. Narody Azii i Afriki 1: 172–186. In Russian (Васильев Л. С. Становление политической администрации (от локальной группы охотников и собирателей к протогосударству – чифдом). Народы Азии и Африки 1: 172–186.
Verencamp S. L. 1983. A Model for the Evolution of Despotic Versus Egalitarian Societies. Anim. Behav. 31: 667–682.
Vermande M. M., and Sterck E. H. 2020. How to Get the Biggest Slice of the Cake. A Comparative View of Social Behaviour and Resource Access in Human Children and Nonhuman Primates. Frontiers in Psychology 11: 2747.
de Waal Frans B. 1987. Tension Regulation and Nonreproductive Function of Sex in Captive Bonobos (Pan paniscus). National Geographic Research 3(3): 318–335.
de Waal Frans B. 1990. Peacemaking among Primates. Cambridge, MA: Harvard University Press.
de Waal Frans B., and Luttrell L. 1989. Toward a Comparative Socioecology of the Genus Macaca: Different Dominance Styles in Rhesus and Stumptail Monkeys. Amer. J. Primatol. 19: 83–109.
de Waal Frans B., and Yoshihara D. 1983. Reconciliation and Redirected Affection in Rhesus Monkeys. Behavior 85: 224–241.
Warner W. L. 1937–1958. A Black Civilization: A Social Study of an Australian Tribe. New York: Harper and Brothers.
Watts D. P. 1992. Social Relationships of Resident and Immigrant Female Mountain Gorillas, I. Male–Female Relationships. Am. J. Primatol. 28: 159–181.
Watts D. P. 1994. Social Relationships of Immigrant and Resident Female Mountain Gorillas. Am. J. Primatol. 32: 13–30.
Watts D. P. 1995. Post-Conflict Social Event in Wild Mountain Gorillas. II. Redirection, Side Direction and Consolation. Ethology 100: 158–174.
Weir S. 2007. A Tribal Order. Politics and Law in the Mountains of Yemen. Austin: University of Texas Press.
White D. R. 1988. Rethinking Polygyny. Current Anthropology 29: 529–558.
White D. R., and Burton M. L. 1988. Causes of Polygyny: Ecology, Economy, Kinship and Warfare. American Anthropologist 90: 871–887.
White I. M. 1978. Aboriginal Women's Status: A Paradox Resolved. Woman's Role in Aboriginal Society, 3rd ed. / Ed. by F. Gale, pp. 36–49. Canberra: Australian Institute of Aboriginal Studies.
Whyte M. K. 1978. Cross-Cultural Codes Dealing with the Relative Status of Women. Ethnology 17: 211–237.
Whyte M. K. 1985. The Relative Status of Women Variables. World Cultures 1(4): stds23–24.dat.
Wilhelm J. H. 1954. Die !Kung-Buschleute. Jahrbuch des Museums für Völkerkunde, Band 12/1953. Leipzig: Otto Harrassowitz.
Williamson D. K., and Dunbar R. 1999. Energetics, Time Budgets and Group Size. Comparative Primate Socioecology / Ed. by P. C. Lee, pp. 320–338. Cambridge: Cambridge University Press.
Woodburn J. C. 1968a. An Introduction to Hadza Ecology. Man the Hunter / Ed. by R. B. Lee, and I. DeVore, pp. 49–55. Chicago, IL: Aldine.
Woodburn J. C. 1968b. Stability and Flexibility in Hadza Residential Groupings. Man the Hunter / Ed. by R. B. Lee, and I. DeVore, pp. 103–110. Chicago, IL: Aldine.
Woodburn J. C. 1972. Ecology, Nomadic Movement and the Composition of the Local Group among Hunters and Gatherers: An East African Example and its Implications. Man, Settlement and Urbanism / Ed. by P. J. Ucko, R. Tringham, and G. W. Dimbleby, pp. 193–206. London: Duckworth.
Woodburn J. C. 1979. Minimal
Politics: The Political Organization of the Hadza of North Tanzania. Politics
in Leadership: A Comparative Perspective / Ed. by
W. A. Shack, and P. S. Cohen. Oxford: Clarendon Press.
Woodburn J. C. 1980. Hunters and Gatherers Today and Reconstruction of the Past. Soviet and Western Anthropology / Ed. by E. Gellner, pp. 95–117. London: Duckworth.
Woodburn J. C. 1982. Egalitarian Societies. Man 17: 431–451.
Woodburn J. C. 1988a. African Hunter-Gatherer Social Organization. Is It Best Understood as a Product of Encapsulation? Hunters and Gatherers. I. History, Evolution and Social Change / Ed. by T. Ingold, D. Riches, and J. Woodburn, pp. 31–64. Oxford: Berg Publishers.
Woodburn J. C. 1988b. Some
Connections between Property, Power and Ideology. Hunters and Gatherers.
II. Property, Power and Ideology / Ed. by T. Ingold,
D. Riches and J. Woodburn, pp. 10–31. Oxford: Berg Publishers.
Wrangham R. W. 1980. An Ecological Model of Female-Bonded Primate Groups. Behavior 75: 262–300.
Wrangham R., and Peterson D. 1996. Demonic Males. Boston – New York: Houghton Mifflin Company.
Yang Martin C. 1945. A Chinese Village. New York: Columbia University Press.
Yengoyan A. A. 1970. Demographic
Factors in Pitjandjara Social Organization. Australian Aboriginal Anthropology
/ Ed. by R. M. Berndt, and C. H. Berndt,
pp. 70–91. Nedlands: University of Western Australia.
Yengoyan
A. A. 1976. Structure, Event, and Ecology in Aboriginal Australia:
A Comparative Viewpoint. Tribes and Boundaries in Australia / Ed. by N. Peterson,
pp. 121–132. Canberra.
Zannella A., Stanyon R., and Palagi E. 2017. Yawning and Social Styles: Different Functions in Tolerant and Despotic Macaques (Macaca Tonkeana and Macaca Fuscata). Journal of Comparative Psychology 131(3): 179.
Zekiye
Eglar. 1957. Punjabi Village Life. Pakistan: Society and Culture
/ Ed. by
S. Maron, pp. 70–75. New Haven: Human Relations Area Files.
Zhao Q.-K. 1993. Sexual Behavior of Tibetan Macaques at Mt. Emei, China. Primates 34: 431–444.
Zhao Q., and Deng Z. 1988. Macaca Thibetana at Mt. Emei, China. II Birth Seasonality. Am. J. Primatol. 16: 261–268.

[1] All Bushmen groups had girls' initiations (accompanied by the Eland dance) which did not envisage any physical endurance trials or surgical operations. Nharo Bushmen (the Central linguistic group), similarly to the !Kung, had both male and female initiations (Silberbauer 1972: 316).
[2] See also White 1978: 37; Keen 1994: 91–96.
[3] It seems necessary to emphasize that, logically speaking, what should be treated as an essential characteristic of the tribal organization is not the conflicts between the residential groups which is absolutely normal as well for the primitive societies lacking any specifically tribal organization (they are considered by Service as belonging to ‘the band level of sociocultural integration’ [Ibid.: 46–98]) but the fact that the tribal organization which puts certain limits to such conflicts makes the feuding parties conflict according to certain rules, provides to the parties highly developed mechanisms of mediation, etc., quite often effectively blocking the most disintegrating consequences of such conflicts but without the alienation of the resident group ‘sovereignty’ (actually Service speaks about it on the pages which follow this quotation, though without the necessary clarity). It also seems necessary to emphasize that the situation, described by Service, may not be necessarily connected only with the complete absence of any supratribal political structures (‘higher authority’) but also with their weakness (which is confirmed in relation to the most tribes of the Middle East); whereas their weakness in many ‘tribal areas’ may often be partly due to the effectiveness of the tribal organization which often allows a sufficiently developed population to live without any strong state structures.
[4] The socio-political organization of the chiefdom is described in a very similar way by most of other political anthropologists: Friedman and Rowlands 1977; Vasil'jev 1980; Earle 1987; Carneiro 1981, 1991, etc.
[5] It is quite hard not to notice that the description of the ‘ideal’ tribal leader by Service resembles rather closely the modern description of the position of the shaykhs among the Yemeni qabâ'il: ‘A shaykh cannot ... make undertakings on his men's behalf simply on the basis of his formal position; each undertaking which affects them must be specifically agreed to...’ (Dresch 1984a: 39). ‘The power which a shaykh may have over groups of tribesmen is not conferred on him by his position. He must constantly intervene in their affairs, and intervene successfully’ (in order to preserve his power) (Ibid.: 41; see also Chelhod 1970, 1979, 1985: 39–54; Dostal 1974, 1990: 47–58, 175–223; Obermeyer 1982; Dresch 1984b, 1989; Abu Ghanim 1985, 1990: 229–251; vom Bruck 1993: 94–95, etc.).
[6] In other words, their diet is closer to the one of the Paleolithic hunter-gatherers than to the one of the intensive preindustrial agriculturists.
[7] Other way out is to seek temporal refuge for the whole band on the territory of the more lucky (in regards to water supply) friendly neighboring band. A similar model was observed for the G/wi of the Central Kalahari Game Reserve (Silberbauer 1972), the !Xõ Bushmen of Southern Kalahari (Heinz 1972). In addition to the yearly rain fluctuations the precipitation regime of Kalahari has 4-year and 10-year cycles of draughts, during which the problem of the re-distribution of the population within the various localities used to become even more relevant (Picaut et al. 1996; Jin 1996: 76–78; Mathhew et al. 1999: 574; Owens and Owens 1985: 59–60, 129–30). A similar situation is described also for the Bushmen of the Karoo regions of South Africa (Cole 1966). On the other side of the globe similar reasons caused the trend toward egalitarianization among the Western Desert Aborigines (so-called Pitjantjara). The precipitation regime here was even more unstable than in the northwestern Kalahari (Leigh and Noble 1969: 74; Tonkinson 1974: 26; Gould 1981: 425). The inhabitants of the drought-stricken areas had to travel up to 300 kilometers in order to reach a refuge (Strehlow 1965: 124–128; Gould 1969: 55–56, 1991; Layton 1986: 18). Gould even described the tactics of the ‘rain hunting’ when Ngatatjara Aborigines moved towards the rain clouds seen at a distance of 80 kilometers (Gould 1977: 21–22; 1981: 430). The situation among the Aranda was different. Here, under the unique for the Central Australia conditions of the relatively well-watered McDonnel Ranges the Aborigines had substantial and quite predictable ecological resources for building elaborated non-egalitarian social structure (Strehlow 1965; Porteus 1931; Spencer and Gillen 1927).
[8] We already mentioned that with respect to the egalitarianism vs. non-egalitarianism correlates, which we considered the only conspicuous exception between the post-industrial and hunter-gatherer egalitarian societies, appears to be Permissiveness on Feeding Sites (supposedly corresponding for the Humans to the presence of privileges in distribution of food). But are the post-industrial egalitarian societies entirely different from the hunter-gatherer egalitarian cultures in economic terms? We do not think so. Indeed, what is the main mechanism of economic egalitarianization among the hunter-gatherers? No doubt, the sharing. Is there a social mechanism in post-industrial egalitarian societies which is similar in its functional role to the ‘primitive’ sharing? And this is simply the social welfare mechanism. You have killed a large animal and you share its meat with those who have not been lucky today. But then you might not be lucky for many weeks and you will get your meat from those who will be lucky. Is the situation in the post-industrial egalitarian societies so much different? You have your job now and you pay your taxes part of which is used to pay unemployment benefits. But then if you lose your job, you will be getting your allowance till you find a new one.