Prebiological Panspermia and the Hypothesis of the Self-Consistent Galaxy Origin of Life

Almanac: Evolution:From Big Bang to Nanorobots
Abstract
We argue that panspermia can mean not only the other place of the origin of life but also another mechanism of the origin of life that increases the probability of the origin of life to many orders of magnitude compared to a single-planet prebiological evolution. The prebiological evolution can be an all-Galaxy coherent process due to the fact that prebiological panspermia and the origin of life are similar to Galaxy-scale second-order phase transition. This mechanism predicts life to have the same chemical base and the same chirality everywhere in the Galaxy.
Keywords: prebiological evolution, panspermia, origin of life, phase transition, Galaxy.
Life was to appear in the process of a natural chemical prebiological evolution. Nobody can estimate now a ‘natural’ duration of the prebiological evolution on a single planet like Earth proceeding from the ‘first principles’ or the experiment. We will show how an independent phenomenological estimation of its time scale can be obtained from the timescale of the Earth biosphere evolution.
Let us consider a number of the first great steps of the biological evolution (hereinafter ‘phase transitions’).
Phase transition 0. The origin of life – about 3.9´109 years ago (Orgel 1998: 91). After the biosphere appeared, it was presented by anucleate anaerobic unicellular organisms – prokaryotes (and, possibly, viruses). Evidently, it existed in such a form without considerable shocks during the first 2–2.5 billion years.
Phase transition 1. The Neoproterozoic revolution (Rozanov 2003: 41). Anaerobic cyanobacteria enriched atmosphere in oxygen which was a strong poison for the anaerobic prokaryotes. This caused an ecological crisis. Apparently, the first one in the history of the Earth. Extinction of the anaerobic prokaryotes started, and the anaerobic prokaryote fauna gave place to the eukaryote and primitive multicellular.
Phase transition 2. The Cambrian explosion (the beginning of the Paleozoic Era) – 570´106 years ago (Carroll 1992, vol. 1: 10). Most modern phylogenetic branches of multicellular organisms (including vertebrates) appeared during tens of millions of years. In the Paleozoic Era the land was gradually inhabited by living creatures. When it was totally inhabited and all corresponding ecological niches were filled, the next evolution crisis occurred.
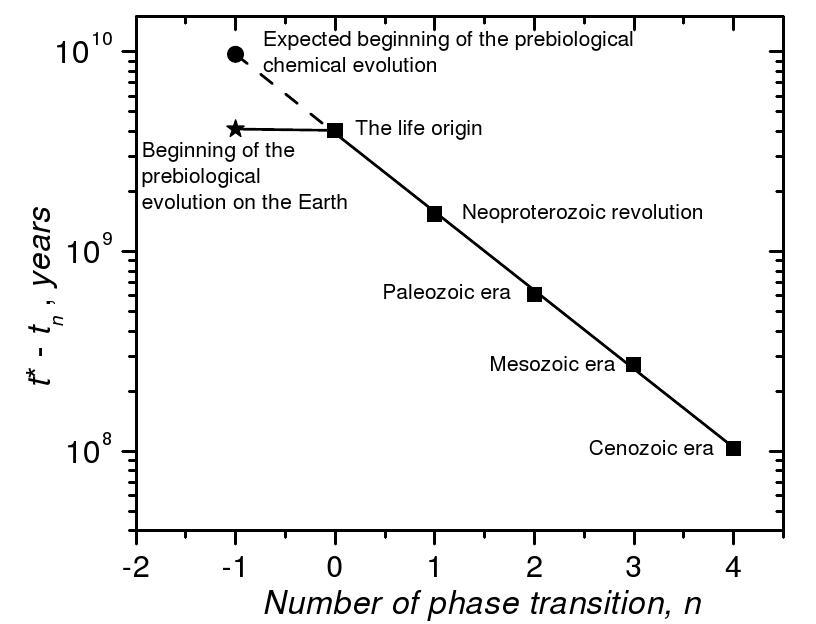
Fig. 1. Extremely short time of the prebiological chemical evolution on the Earth produces a ‘hockey’ stick anomaly in the exponential scale of time of the evolution
Phase transition 3. The revolution of reptiles (the beginning of the Mesozoic Era) – 235´106 years ago (Carrol 1992: V. 1, V. 2). Practically all species of Paleozoic amphibia die out. Reptiles become leaders of evolution on land.
Phase transition 4. The revolution of mammals (the beginning of the Cenozoic Era) – 66´106 years ago (Carrol 1992: V. 2, V. 3). Dinosaurs die out. Mammals and birds become the leaders of evolution on land.
It is not difficult to see that the duration of the phases of biosphere evolution steadily decreases from the past to the present. Furthermore, the sequence of durations of the phase transitions forms a geometric series T0/an with a»2.7 in good approximation (and the limit point t* of the series fits very good the present moment of time – we are living near the point of the singularity of evolution [Panov 2005: 220–225]).
We see that the higher is the organization level of the biosphere, the higher is the evolution rate. Since any prebiological system has a lower organization level than the biological one, then it seems that the prebiological evolution rate must be even lower than the rate of the subsequent evolution of the biosphere. Furthermore, one can speculate that the duration of the prebiological evolution belongs to the same geometric series of the phases of the biosphere evolu- tion and estimate expected duration of the prebiological evolution by its extrapolation back in time. It is clear that this is only an incomplete induction; our speculations are not a proof; this estimation should be considered as a conjecture.
Using the duration of the first step of the biological evolution 3.9´109 – 1.5´109 = 2.4´109 years, we get an estimation of the duration of the last phase of the prebiological chemical evolution to be τchem = 2.4´109 ´ 2.7 = 6.4´109 years. This is the lower limit of the total duration of the prebiological evolution since the latter can consist of many phases.
The value τchem » 6´109 years is very large. At the same time, there is evidence that the duration of the prebiological chemical evolution on the Earth did not exceed 0.2´109 years (Orgel 1998: 91). An obvious contradiction is present and this contradiction is clearly seen in Fig. 1. It can be solved in the following way. The duration of the ‘natural’ prebiological chemical evolution actually is of order of 6 billion years (or even more), but it occurred not on the Earth, but on the other planets near stars that are much older than the Sun. Life appeared on the Earth as a result of the process of interstellar panspermia from these old planets. However, if the biological panspermia took place, then the prebiolo-gical panspermia could be quite possible as well. The products of the pre-biological chemical evolution must be less sensitive to difficulties of cosmic missions (hard radiation, cold and vacuum) than any biological systems. What is a typical time scale of expansion of a prebiological or biological ‘infection’ over the Galaxy?
Let us refine some details of the panspermia mechanism. Suppose the question is on expansion of a biological or prebiological product characterized by a high elasticity and competitiveness. Upon getting to a planet suitable for adaptation, such a product must expand over the planet surface in some thousands of years or even faster, replacing local weaker systems. As a result, the planet itself becomes a source of panspermia of this advanced product of evolution. If its host star flies near another star, then the latter can be infected and become an object of panspermia too. Then the spread of the product of evolution would have not the diffusion character, but the character of an autowave propagating at a constant velocity, approximately as it occurs in epidemics. The typical velocity of peculiar chaotic motion of stars is decisive. Its value – about 30 km per second – is the typical velocity of the panspermia wave in the Galaxy. To model it, the pure Huygens principle can be used. Of course, the model contains a lot of simplifications. So, for instance, the typical peculiar velocities can differ at different distances from the Galaxy center, etc. But the model is suitable to make a rough estimate of the time scale of the process.
Fig. 2 shows the results of digital simulation of panspermia wave propagation in the Galaxy fulfilled with the above assumptions taking into account differential rotation of the galactic disk. It can be seen from Fig. 2 that due to this rotation the process is practically finished in two galactic years (one galactic year – the period of rotation of the Sun around the Galaxy center – is equal to 216 million years), and 70 per cent of the Galaxy volume is inhabited for about 300 million years.
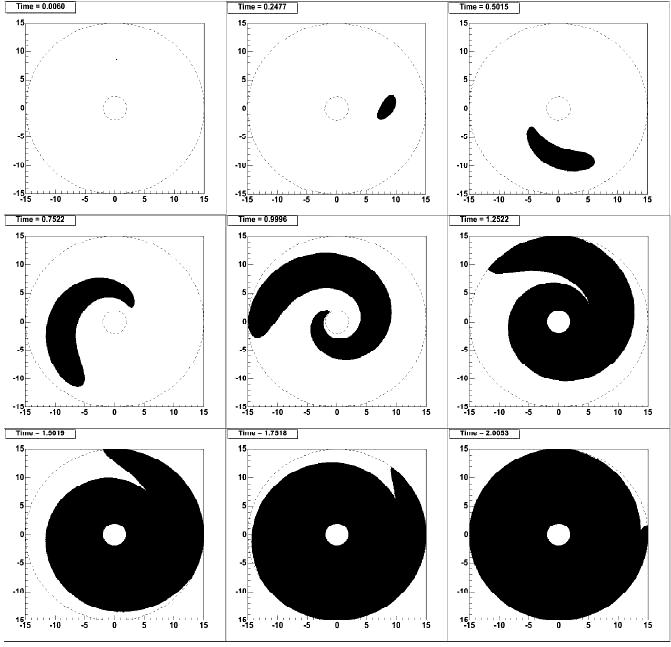
Fig. 2. Digital model of propagation of panspermia wave in the Galaxy disc. Time in figures is shown in galactic years. The subsequent times of the phases are (from upper-left to top-down): 0.0060, 0.2477, 0.5015, 0.7522, 0.9996, 1.2522, 1.5019, 1.7518, 2.0053. The Galaxy rotates clockwise
So, we get two time scales: one long scale of τchem ≈ 6´109 years (or more), this is a scale of natural duration of the prebiological chemical evolution on an isolated planet; the other short scale of τpansp ≈ 0.3´109 years is a scale of duration of the process of galactic panspermia. From the two very different time scales it follows that the prebiological chemical evolution on separate planets could not occur independently of processes on other planets.
Let us suppose that a stable and competitive prebiological system appears on a planet at the prebiological evolution stage of the Galaxy (i.e. before life appeared for the first time). This is quite a random event. Then, during a short time, of order τpansp, this prebiological system spreads over the whole volume of the Galaxy displacing less effective local prebiological systems because of the ordinary natural selection. This is a mechanism of natural selection at the prebiological level on a scale of the whole Galaxy. Due to the condition τpansp << τchem, this process must synchronize (with an accuracy of τраnsp) the prebiological evolution in the whole volume of the Galaxy. As a result, life originates almost simultaneously on all planets having suitable conditions for its existence, with one molecular basis (in terms of the basis of genetic code, etc.) and with one chirality. This event resembles the non-equilibrium phase transition of second order. Thus, the prebiological chemical evolution and the origin of life can be a self-consistent collective process, but not a process located on separate planets as is usually supposed.
If the mechanism of the self-consistent Galaxy origin of life operated, then a gigantic burst of inhabitance of planets with life must have taken place in the Galaxy soon after life appeared somewhere for the first time. After that life could not arise anywhere in the process of the natural prebiological evolution since the natural prebiological process cannot compete with much faster processes of panspermia.
It is widely believed that the probability of self-generating life on any separate planet is vanishingly small. For instance, the origin of life on an isolated the Earth-type planet with suitable conditions can take a billion of billions years or some so absurdly long time. If the prebiological evolution proceeded independently on different planets, then, at present, life would not exist at all or would be a quite unique phenomenon. However, if an effective process of the prebiological panspermia is possible, then any random success of the prebiological evolution on one of about 109 planets of the Galaxy becomes the property of other planets practically immediately. In other words, the probability of such an event on any separate planet increases 109 times! And the rate of the prebiological evolution increases as well. Thus, even if the self-generating origin of life is practically improbable under the conditions of an isolated planet, it can be quite probable because of prebiological panspermia (the last idea is taken from G. A. Skorobogatov, 2004, private communication). Panspermia provides other place of the origin of life of course, but it also does provide other mechanism and other way of the origin.
References
Carroll R. L. 1992. Vertebrate Paleontology and Evolution. 3 vols. Moscow: Mir. In Russian (Кэрролл Р. Палеонтология и эволюция позвоночных. B 3 т. М.: Мир).
Orgel L. E. 1998. The Origin of Life – How Long did it Take? Origins of Life and Evolution of the Biosphere 28(1): 91–96.
Panov A. D. 2005. Scaling Law of the Biological Evolution and the Hypothesis of the Self-Consistent Galaxy Origin of Life. Advances in Space Research 36: 220–225.
Rozanov A. Yu. 2003. Fossil Bacteria, Sedimentogenesis and Early Stages of Evolution of the Biosphere. Paleontologicheskiy zhurnal 6: 41–49. In Russian (Розанов А. Ю. Ископаемые бактерии, седиментогенез и ранние стадии эволюции биосферы. Палеонтологический журнал 6: 41–49).